
 Open Access
Open Access Abstract
The fresh green algal Haematococcus pluvialis Flotow was proved to be the starting material for the production of biofuel, high lipid content along with astaxanthin, a high value colorant. In this study, lipid accumulation in H. pluvialis cultured in liquid Bold’s Basal medium aerated was investigated for a period of 12 weeks. Lipid accumulation was evaluated through the expression of two genes: BC (biotin carboxylase, initial gene) and FATA (acyl-acyl carrier protein thioesterase, end gene) in the process of fatty acid biosynthesis with Real-time RT-PCR, lipid determination by Nile Red and biodiesel quantifying by transesterification. The results showed that the expression of two BC and FATA genes was recorded at all weeks of culture. However, the expression of BC and FATA genes increased gradually from the week 9 (1.3, 4.1, respectively) to week 11 (1.7, 30.9, respectively). Meanwhile, yellow fluorescence in the microalgal cells showed that lipid appeared from week 6 to week 12. The obtained biodiesel increased slowly from week 8 (0.036 mg/mL) to week 12 (0.041 mg/mL). At week 11, the expression values of both BC gene (1.7) and FATA gene (30.9) were maximized, leading to the highest biodiesel content at the week 12.
ĐẶT VẤN ĐỀ
Việc hướng tới sản xuất nhiên liệu sinh học (biofuel), một loại nhiên liệu tái tạo được sản xuất từ sinh khối sinh học được xem là một giải pháp cho việc thay thế nhiên liệu truyền thống đang cạn kiệt 1 . Việc sản xuất năng lượng sinh học chủ yếu dựa trên các nguyên liệu khởi đầu có hàm lượng acid béo cao 2 , 3 . Các công trình nghiên cứu trên thế giới đã chứng minh rằng tảo lục nước ngọt Haematococcus pluviali s Flotow có thể được sử dụng để sản xuất nhiên liệu sinh học thế hệ thứ ba 1 , 4 . Đặc biệt, trong nghiên cứu của Lei và cộng sự (2012) 1 , con đường sinh tổng hợp acid béo tự do trong H. pluvialis đã được công bố với sự tham gia của nhiều gen khác nhau nhằm xúc tác chuyển đổi acetyl-CoA thành acid béo tự do, trong đó, gen biotin carboxylase ( B C, EF523480) và acyl-acyl carrier protein thioesteras e ( FAT A , HM560034) là hai gen đóng vai trò quan trọng trong giai đoạn khởi đầu và kết thúc của quá trình sinh tổng hợp acid béo. Gen BC mã hóa cho biotin carboxylase, là một trong ba tiểu phần của acetyl-CoA carboxylase biotin carboxylase, xúc tác chuyển nhóm carboxyl từ acetyl-CoA hình thành malonyl-CoA khởi đầu cho quá trình sinh tổng hợp acid béo 1 , 5 , 6 . Với FATA , gen này mã hóa cho enzyme fatty acyl-ACP thioesterase, xúc tác chuyển đổi malonyl-ACP thành acid béo tích lũy trong H. pluvialis 1 , 7 .
Trong nghiên cứu trước đây, chúng tôi đã thành công trong việc nuôi cấy vi tảo và thiết kế cặp mồi và xây dựng quy trình PCR khuếch đại sự hiện diện của các gen BC và FATA 8 , 9 . Nghiên cứu tiếp theo này, sự biểu hiện của hai gen BC và FATA trên vi tảo H. pluvialis được theo dõi trong quá trình nuôi vi tảo trong môi trường lỏng Bold ’s Basal (BB) được sục khí ở các tuần khác nhau, nhằm tìm hiểu mối liên quan giữa sư ̣biểu hiện của các gen nêu trên với sự tích lũy lipid ở tảo H. pluvialis .
VẬT LIỆU VÀ PHƯƠNG PHÁP
Vật liệu
Vi tảo H. pluvialis Flotow được phòng Sinh học thực nghiệm của Viện Nghiên cứu và Nuôi trồng Thủy sản II, TP. Hồ Chí Minh cung cấp lại từ bộ sưu tập giống tảo của phòng Công nghệ Tảo, Viện Công nghệ Sinh học thuộc Viện Hàn lâm Khoa học và Công nghệ Việt Nam, Hà Nội.
Vi tảo sau khi thu nhận được tiến hành nuôi cấy trong 250 mL môi trường lỏng BB được sục khí. Sau 9 tuần nuôi cấy, 50 mL môi trường chứa vi tảo được thu nhận và chuyển sang bình sục khí 500 mL có chứa 250 mL môi trường lỏng BB ở pH 7, nhiệt độ 25 ± 3 °C, cường độ ánh sáng 50 µmol photon m -2 s -1 và chu kỳ chiếu sáng là 12 giờ/ngày. Sự tăng trưởng và tích lũy lipid của vi tảo được tiến hành đánh giá trong thời gian 12 tuần (Quy ước: tuần 1 đến tuần 12).
Phương pháp
Kiểm tra sự hiện diện lipid bằng thuốc nhuộm Nile Red dưới kính hiển vi huỳnh quang
200 µL vi tảo từ bình chứa môi trường lỏng BB được sục khí ở các thời gian nuôi cấy khác nhau (từ tuần 0 đến tuần 12) thu được theo phương pháp xác định trọng lượng tươi, được nhuộm với 5 µL thuốc thử Nile Red (pha trong DMSO 0,5 mg/mL). Ủ trong tối 10 phút, và quan sát huỳnh quang màu vàng dưới kính hiển vi huỳnh quang với nguồn ánh sáng xanh với bước sóng 460 - 480 nm 10 .
Xác định hàm lượng dầu sinh học tích lũy ở vi tảo H. pluvialis
1 mL môi trường chứa vi tảo H. pluvialis từ bình chứa môi trường lỏng BB được sục khí ở các thời gian nuôi cấy khác nhau được dùng để thu sinh khối theo phương pháp xác định trọng lượng tươi. Sau đó bổ sung 12 mL chloroform và 24 mL methanol vào sinh khối thu được. Tiếp theo ly tâm dung dịch thu được ở 3.500 vòng/phút trong 15 phút. Sau ly tâm, thu dịch nổi và bổ sung thêm 6,8 mL methanol, 1,2 mL NaOH 0,1 N và 8 mL chloroform. Tiến hành ủ cách thủy dung dịch thu được ở 90 o C trong 40 phút và sau đó để nguội. Tiếp theo thêm vào 4 mL nước cất để dung dịch phân lớp. Lớp dưới cùng là dầu sinh học 11 . Với phương pháp này, cả lipid tích lũy trong tế bào lẫn lipid thoát ra môi trường nuôi cấy đều được thu nhận bằng sự ester hóa các acid béo.
Tách chiết mRNA tổng số
10 mg vi tảo H. pluvialis được thu nhận ở các tuần nuôi cấy khác nhau. Sau khi thu cặn, 900 µL TRIzol® và 200 µL chloroform được bổ sung để ly giải tế bào. Sau đó, tiến hành ly tâm mẫu ở 13.000 vòng/phút trong 10 phút, thu pha nổi và bổ sung 0,2 ml chloroform. Sau đó, tiếp tục tiến hành ly tâm ở điều kiện 13.000 vòng/phút trong 10 phút, thu pha nổi, bổ sung thêm isopropanol theo tỉ lệ 1:1, ủ trong 3 giờ ở -20 o C. Ly tâm ở tốc độ 13.000 vòng / phút, trong 10 phút. Loại bỏ pha nổi, thêm vào cặn 1 mL ethanol 70 %, lắc đều. Ly tâm 13.000 vòng/phút, trong 10 phút. Loại bỏ pha nổi, để khô tự nhiên, thêm vào 50 µL DEPC (diethyl pyrocarbonate) và trữ mRNA ở -20 o C để sử dụng cho các thí nghiệm sau 12 .
Phân tích sự biểu hiện của gen BC và FATA bằng phương pháp Real-time RT-PCR
15 µL RNA tổng số được sử dụng để thực hiện phản ứng Reversed transcriptase PCR bằng kit cDNA synthesis kit (Thermo Scientific) với cặp mồi Random được cung cấp trong bộ kit. cDNA sau khi thu nhận được tiến hành đo mật độ quang và lưu trữ ở nhiệt độ -20 o C cho việc phân tích sự biểu hiện các gen mục tiêu sau này.
Kỹ thuật Real-time RT-PCR được sử dụng để khảo sát sự biểu hiện của hai gen BC và FATA ở các tuần nuôi cấy khác nhau, mỗi thí nghiệm được lặp lại ba lần (Máy sử dụng: MyGo Pro real-time PCR). Gen beta-actin ( β-acti n) được sử dụng làm chứng nội cho phản ứng này. Thành phần phản ứng bao gồm: 5 µl cDNA của H. pluvialis được bổ sung với 0,5 µL mồi xuôi, 0,5 µL mồi ngược, 9 µL nước không chứa nuclease, 5 µL SYBR I (Bioline). Chu kỳ phản ứng được thiết lập như sau: 95 o C trong 60 giây, 40 chu kỳ (95 o C trong 30 giây, 40 o C trong 30 giây). Chứng âm có thành phần tương tự nhưng thay thế cDNA bằng nước không chứa nuclease. Bảng mồi sử dụng trong nghiên cứu này được thể hiện ở Table 1 . Tính chất biểu hiện của các gen BC và FATA ở các tuần khác nhau được tính toán thông qua giá trị định lượng tương đối 2 -∆∆Ct . Tất cả các giá trị Ct (Cycle threshold) để định lượng sự biểu hiện của mRNA các gen BC và FATA được quy chiếu với mRNA chứng nội là gen β -actin thành giá trị ΔCt.
| Gen đích | Mồi | Trình tự 5’ – 3’ |
| BC | xuôi: BC -F | GAAGGTGATGATCGCCAACC |
| ngược: BC -R | TGGACGTGCAGCGAGTTCT | |
| FATA | xuôi: FATA -F | AGACTCGTTCAGCGAGGAGC |
| ngược: FATA -R | CATGCCCACAGCATGGTTCCC | |
| Beta-actin | xuôi: ACT -F ngược: ACT -R | ACCTCAGCGTTCAGCCTTGT TGGTCCACGACACCATCAAC |
KẾT QUẢ
Sự biểu diện của gen BC và FATA theo thời gian nuôi vi tảo
Phương pháp Real-time RT - PCR được sử dụng để khảo sát sự biểu hiện mRNA hai gen BC và FATA ở các tuần nuôi cấy, ghi nhận các mRNA biểu hiện ở tất cả các tuần nuôi cấy, từ tuần thứ 1 đến tuần thứ 12 ( Figure 1 ).
Figure 1 . Kết quả Real-time RT-PCR từ mRNA của (A) gen BC , (B) gen FATA của vi tảo được nuôi cấy trong bình sục khí có chứa môi trường lỏng BB theo thời gian từ tuần 1 - 12.
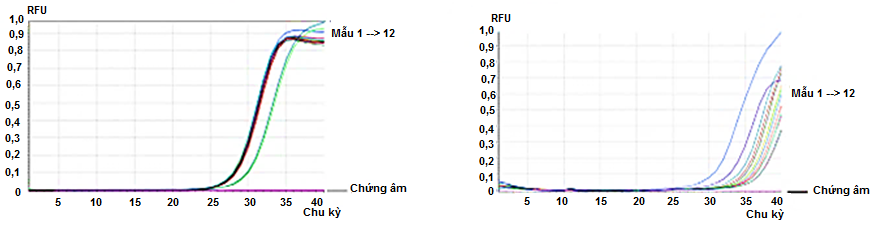
Giá trị định lượng tương đối (2 -∆∆Ct ) đánh giá sự biểu hiện của BC và FATA được tính toán và so sánh với mốc biểu hiện của hai gen này thông qua trị số trung bình của các tuần từ thứ 2 đến thứ 5. Kết quả ghi nhận ở Table 2 cho thấy : biểu hiện của gen BC và FATA bắt đầu tăng ở tuần 6, gen BC và FATA lần lượt tăng biểu hiện gấp 1,3 và 6,3 lần so với trị số trung bình của các tuần 2 - 5. Tuy nhiên, sự tăng này chỉ thật sự ổn định từ tuần thứ 9, 10 và đạt cực đại ở tuần thứ 11. Từ tuần thứ 12, biểu hiện của hai gen đều giảm ( Table 2 ).
| Tuần | Gen beta-actin | Gen BC | Gen FATA | ||
| Ct | Ct | 2-ΔΔCt | Ct | 2-ΔΔCt | |
| 1 | 28,1 | 27,2 | - | 30,1 | - |
| 2 - 5 | 27,4 | 28,1 | 1,0 | 35,8 | 1 |
| 6 | 27,6 | 27,4 | 1,3 | 35,0 | 6,3 |
| 7 | 27,2 | 27,6 | 0,9 | 34,4 | 4,8 |
| 8 | 27,4 | 27,2 | 0,7 | 35,9 | 0,7 |
| 9 | 27,4 | 27,4 | 1,3 | 35,3 | 4,1 |
| 10 | 27,5 | 27,4 | 1,4 | 34,1 | 19,0 |
| 11 | 29,0 | 27,5 | 1,7 | 32,4 | 30,9 |
| 12 | 28,1 | 29,0 | 0,8 | 33,6 | 10,9 |
Sự tích lũy lipid theo thời gian nuôi vi tảo
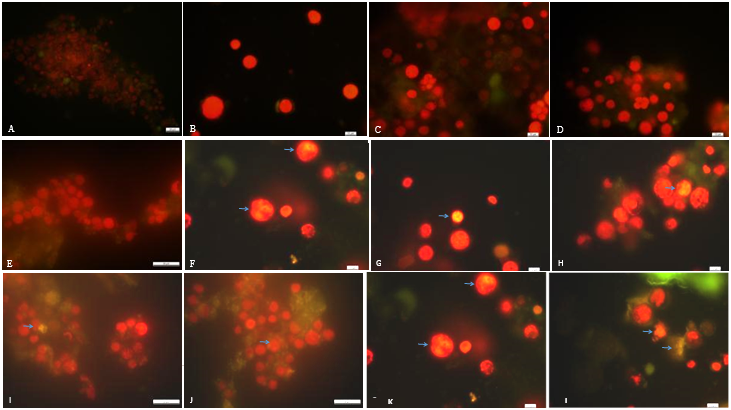
Các tế bào vi tảo được tiến hành nhuộm với Nile Red và quan sát tín hiệu huỳnh quang, cho kết quả: từ tuần 1 đến tuần 5, không quan sát được sự phát huỳnh quang ở vi tảo ( Figure 2 A E ) ; Từ tuần 6, bắt đầu quan sát được sự phát huỳnh quang ở tế bào vi tả o và tín hiệu huỳnh quang tiếp tục được ghi nhận đến tuần 8 ( Figure 2 F -H ). Từ tuần 9 đến tuần 12, tín hiệu huỳnh quang được ghi nhận xuất hiện nhiều bên ngoài tế bào vi tảo ( Figure 2 I - L ).
Figure 2 . Tích lũy lipid ở vi tảo quan sát qua các tuần bằng phương pháp nhuộm Nile Red. Chú thích: A - L: lần lượt tương ứng với các tuần 1 đến tuần 12. Dấu mũi tên chỉ các tế bào vi tảo phát huỳnh quang có chứa lipid. Thanh ngang 10 µm.
Hàm lượng dầu sinh học theo thời gian nuôi vi tảo
Dầu sinh học thu nhận được từ nuôi cấy vi tảo trong môi trường lỏng BB sục khí theo thời gian được ghi nhận tăng chậm từ tuần 8 đến tuần 12. Hàm lượng dầu cao nhất được ghi nhận ở tuần thứ 12, đạt 0,041 ± 0,007 mg/mL ( Table 3 ).
| Thời gian nuôi cấy (tuần) | Dầu sinh học tổng cộng (mg/mL) |
| 1 → 7 | 0,0 ± 0,00 |
| 8 | 0,036 ± 0,003ab |
| 9 | 0,032 ± 0,004ab |
| 10 | 0,027 ± 0,002b |
| 11 | 0,032 ± 0,010b |
| 12 | 0,041 ± 0,007a |
THẢO LUẬN
Trong những thập niên gần đây, hướng tới việc giải quyết sự khủng hoảng nguồn nhiên liệu đang cạn dần, nhiên liệu sinh học được xem là một giải pháp thay thế cho nhiên liệu truyền thống đang cạn kiệt dần. Việc sử dụng vi tảo làm nguyên liệu sản xuất nhiên liệu sinh học được xem là một phương pháp khả thi do nhiều đặc tính ưu việc khác nhau, chẳng hạn việc nuôi trồng vi tảo không cạnh tranh đất sản xuất nông nghiệp, không làm ô nhiễm môi trường, thích nghi trong nhiều điều kiện sống khác nhau, quá trình trao đổi chất lớn. Trong đó, vi tảo nước ngọt Haematococcus pluvialis được chứng minh có khả năng sản xuất nhiều dầu sinh học và astaxanthin có giá trị kinh tế cao 13 . Do đó, trong nghiên cứu này chúng tôi đánh giá sự tích lũy lipid ở vi tảo H. pluvialis qua các tuần theo thời gian nuôi. Bằng phương pháp nhuộm Nile Red và quan sát dưới hiển vi huỳnh quang, sự tích lũy lipid ở vi tảo được đánh giá một cách định tính không ghi nhận được lipid ở vi tảo H. pluvialis trong 5 tuần đầu khi vi tảo được nuôi trong môi trường lỏng BB, sục khí. Xét về sự tăng trưởng của vi tảo, giai đoạn chuẩn bị tăng trưởng của vi tảo là từ tuần 0 - 1, giai đoạn tăng mật độ tế bào là từ tuần 2 - 6; trong đó, tuần cuối cùng của giai đoạn tăng mật độ tế bào, lipid tích lũy mới được ghi nhận bằng phương pháp nhuộm Nile Red. Kết quả nghiên cứu này phù hợp với nghiên cứu của Borowitzka và Moheimami (2013): Khi vi tảo tăng trưởng chậm và không cần màng mới, tế bào chuyển sang tổng hợp TAG và chờ điều kiện thích hợp để tiếp tục tăng trưởng 14 . Trong khi đó, bằng phương pháp đánh giá sự biểu hiện gen, các gen thiết yếu trong con đường sinh tổng hợp lipd đã được ghi nhận từ các tuần 1 - 5 và được đánh giá có tăng từ tuần 6. Việc đánh giá tính chất biểu hiện của mRNA BC và FATA được tính toán thông qua giá trị 2 -∆∆Ct với chứng nội được sử dụng là gen beta-actin . Gen beta-actin là một gen giữ nhà (house keeping gene) có tính bảo tồn cao và luôn được biểu hiện ở tất cả các tế bào Eukaryote. Kết quả cho thấy, trong hai tuần 7 - 8, tức là vi tảo thuộc giai đoạn tăng trưởng nhanh, sự biểu hiện các gen BC và FATA không tăng, có giảm nhẹ so với tuần thứ 6, sự biểu hiện các gen này chỉ thực sự gia tăng và ổn định vào tuần thứ 9, 10 và đạt cực đại ở tuần thứ 11 (gen BC và FATA tăng biểu hiện lần lượt 1,7 và 30,9 lần so với trị số trung bình của các tuần 1 - 5). Kết quả này phù hợp với kết quả đánh giá định tính sự tích lũy lipid bằng thuốc nhuộm Nile Red: lipid được ghi nhận bên trong tế bào vi tảo trong suốt giai đoạn tăng trưởng nhanh (tuần 7 - 8) của tế bào vi tảo. Tuy nhiên, hàm lượng lipid chưa đủ cao để có thể chuyển hóa ester tạo dầu sinh học ở các tuần này. Ở các tuần 9 - 12, tức là giai đoạn vi tảo có tăng trưởng chậm, lipid tích lũy được xuất ra bên ngoài tế bào, ở tuần thứ 12, lúc này vi tảo bước vào giai đoạn cuối của qua trình tăng trưởng chậm, hàm lượng dầu sinh học được ghi nhận cao nhất (0,041 ± 0,007 mg/mL), trong khi sự biểu hiện của cả hai gen BC và FATA đã bắt đầu giảm (gen BC và FATA giảm biểu hiện lần lượt 0,8 và 10,9 lần so với trị số trung bình của các tuần 1 - 5). Ở tuần 12, vi tảo có tích lũy lipid cao nhất nên cần thu nhận sinh khối để tách dầu sinh học.
KẾT LUẬN
Trong nghiên cứu này, chúng tôi đã thành công trong việc phân tích sự tích lũy lipid ở vi tảo H. pluvialis trong môi trường lỏng BB trong thời gian 12 tuần nuôi cấy. Kết quả phân tích biểu hiện gen BC và FATA , cũng như định tính lipid ghi nhận lipid tích lũy trong vi tảo từ tuần thứ 6. Dầu sinh học được ghi nhận từ tuần 8 và đạt cực đại ở tuần 12.
DANH MỤC TỪ VIẾT TẮT
BB : Bold’s Basal
BC : biotin carboxylase
FATA : acyl-acyl carrier protein thioesterase
RT-PCR : Reverse Transcription - Polymerase Chain Reaction
DMSO : Dimethyl sulfoxide
DEPC : Diethyl pyrocarbonate
mRNA : messenger Ribonucleic acid
RFU : Relative fluorescence units
RNA : Ribonucleic acid
cDNA : complementary Deoxyribonucleic acid
Ct : Cycle threshold
TAG : Triacylglycerol
XUNG ĐỘT LỢI ÍCH
Nhóm tác giả không có bất kỳ xung đột lợi ích lẫn nhau.
ĐÓNG GÓP CỦA TÁC GIẢ
Nguyễn Trần Đông Phương: thực hiện thí nghiệm, thu thập và xử lý các dữ liệu thu được.
Lê Huyền Ái Thúy, Bùi Trang Việt: đóng góp chính trong việc viết và chỉnh sửa bản thảo.
References
- Lei A, Chen H, Shen G, Hu Z, Chen L, Wang J. Expression of fatty acid synthesis genes and fatty acid accumulation in Haematococcus pluvialis under different stressors. Biotechnol Biofuels. 2012;5(18):2-11. Google Scholar
- Vicente G, Martínez M, Aracil J. Optimisation of integrated biodiesel production. Part I. A study of the biodiesel purity and yield. Bioresour Technol. 2007;98:1724-1733. Google Scholar
- Zhou Y J, Buijs N A, Siewers V, Nielsen J. Fatty Acid-Derived Biofuels and Chemicals Production in Saccharomyces cerevisiae. Front Bioeng Biotechnol. 2014;2:32. Google Scholar
- Razon Luis F, Tan Raymond R. Net energy analysis of the production of biodiesel and biogas from the microalgae: Haematococcus pluvialis and Nannochloropsis. Applied Energy. 2011;88:3507-3514. Google Scholar
- Ohlrogge J, Browse J. Lipid biosynthesis. Plant Cell. 1995;7:957-970. Google Scholar
- Janiyani K, Bordelon T, Waldrop G L, Cronan J E Jr. Function of Escherichia coli biotin carboxylase requires catalytic activity of both subunits of the homodimer. J Biol Chem. 2001;276:29864-29870. Google Scholar
- Jones A, Davies H M, Voelker T A. Palmitoyl-acyl carrier protein (ACP) thioesterase and the evolutionary origin of plant acyl-ACP thioesterases. Plant Cell. 1995;7:359-371. Google Scholar
- Phương Nguyn Trần Đông, Thúy Lê Huyền Ái, Việt Bùi Trang. Ảnh hưởng cùa các chất điều hòa sinh trưởng thực vật lên sự sinh trưởng của vi tảo Haematococcus pluvialis Flotow. Tạp chí Công nghệ sinh học. 2015;13(2):269-247. Google Scholar
- Phuong Nguyen Tran Dong, Thuan Lao Duc, Thuy Le Huyen Ai, Viet Bui Trang. Initial studies on Biotin carboxylase (BC) and acyl-acyl carrier protein thioesterase (FATA) genes in Haematococcus pluvialis Flotow. J Biotech. 2016;14(1A):531-538. Google Scholar
- Gwak Y, Hwang Y S, Wang B, Kim M, Jeong J, Lee C-G, Hu Q, Han D, E Jin. Comparative analyses of lipidomes and transcriptomes reveal a concerted action of multiple defensive systems against photooxidative stress in Haematococcus pluvialis. Journal of Experimental Botany. 2014;65(15):4317-4334. Google Scholar
- Johnson M B, Wen Z. Preparation of biodiesel fuel from the microalga Schizochytrium limacinum by direct transesterification of algal biomass. Energy and Fuels, In Progress. 2009;:. Google Scholar
- Macedo N J, Ferreira T. Maximizing total RNA yield from TRIzol® reagent protocol: a feasibility study. UB-ASEE. 2014;(1):1-7. Google Scholar
- Razon L F, Tan R R. Net energy analysis of the production of biodiesel and biogas from the microalgae: Haematococcus pluvialis and Nannochloropsis. Applied Energy. 2011;88:3507-3514. Google Scholar
- Borowitzka M A, Moheimani N R. Energy from microalgae: a short story. Algae for biofuels and Energy. 2013;:1-15. Google Scholar





