
 Open Access
Open Access Abstract
In the finding of new antimicrobial agents to respond to drug-resistant bacterial pathogens, the genus Streptomyces is still an important source of secondary metabolites associated with antibacterial activity. Strikingly, two-thirds of antibiotics used in human medicine are secondary metabolites from Streptomyces bacteria, among which the biosynthetic pathway of several antibiotics was studied. We report here the isolation of an endophytic bacterial strain from Bougainvillea glabra collected in Vung Tau City, Vietnam. This bacterial strain showed strong antibacterial activity against multiple pathogenic bacteria, including those belonging to the ESKAPE pathogens group. Cultural and molecular characterization identified the isolated strain as Streptomyces sp. SS200. A DNA transfer system by bacterial conjugation was established in Streptomyces sp. SS200 to pave the way for following studies on biosynthetic pathways of secondary metabolites having antibacterial activity in this strain. The key conditions in the conjugal transfer were investigated, including the recipient’s cellular forms, donor Escherichia coli strains, effects of cation2+, conjugation media, and antibiotics overlaying time. The mycelial form of Streptomyces sp. SS200 gave a 5-fold higher conjugation frequency than the spore form. ET12567 strain carrying the mobilizing plasmid pUZ8002 had the highest frequency of conjugation among two E. coli donors, specifically at a double frequency than SM10pir strain. Ca2+ gave more exconjugants than Mg2+, and the highest frequency of conjugation was obtained at 10 mM of this cation. Of four conjugation media, the exconjugants between E. coli and Streptomyces were only found on the Soya Flour Mannitol medium. The overlaying time of antibiotics was optimal at 24 hours of conjugation. These conjugal conditions will be used for the following experiments to find gene candidates involved in the biosynthesis of secondary metabolites with antibacterial activity in Streptomyces sp. SS200.
MỞ ĐẦU
Vi khuẩn gây bệnh liên quan đến kháng kháng sinh hiện vẫn là mối đe dọa sức khoẻ cộng đồng mang tính toàn cầu. Trong đó, các vi khuẩn thuộc nhóm ESKAPE bao gồm Enterococcus faecium , Staphylococcus aureus , Klebsiella pneumoniae , Acinetobacter baumannii , Pseudomonas aeruginosa và Enterobacter spp được Tổ chức Y tế Thế giới (WHO) xác định là các chủng gây bệnh nguy hiểm do khả năng tồn tại của chúng trong môi trường bệnh viện do có khả năng tạo ra biofilm và nhất là khả năng kháng lại các kháng sinh điều trị của chúng 1 . Hiện nay, số ca tử vong do các bệnh nhiễm khuẩn ước tính khoảng 700.000 ca mỗi năm trên toàn thế giới 2 . Tuy nhiên, tình trạng này càng trở nên phức tạp và nan giải hơn khi số lượng kháng sinh mới được đưa vào điều trị lâm sàng giảm dần từ những năm 70 của thế kỷ 20 cho đến hiện nay với một ví dụ là chỉ có bảy kháng sinh mới được đưa vào điều trị lâm sàng trong giai đoạn 2013-2017 3 . Nếu tình trạng thiếu thuốc kháng sinh tiếp tục diễn ra, số bệnh nhân tử vong do nhiễm trùng do vi khuẩn sẽ lên đến con số mười triệu ca mỗi năm vào năm 2050 3 .
Để giải quyết vấn đề thiếu kháng sinh trong điều trị bệnh nhiễm khuẩn, đã có nhiều nghiên cứu tìm kiếm các kháng sinh mới từ các nguồn khác nhau như thực vật, vi sinh vật hoặc tổng hợp hóa học. Trong số các nguồn kể trên, vi sinh vật đóng một vai trò quan trọng trong việc tìm kiếm các chất kháng khuẩn mới, trong đó Streptomyces là các vi khuẩn sản xuất kháng sinh nổi trội. Đáng chú ý là các vi khuẩn Streptomyces là các vi khuẩn sản xuất hơn hai phần ba số kháng sinh được biết cho đến nay 4 . Vì vậy, nhóm vi khuẩn này vẫn là đối tượng quan trọng để phân lập các kháng sinh mới mặc dù chúng đã được khai thác từ thập niên 40 của thế kỷ 20 cho mục tiêu này.
Trong số các nguồn tự nhiên để phân lập vi sinh vật có hoạt tính kháng khuẩn, vi sinh vật nội sinh gần đây được tập trung nghiên cứu vì chúng chưa được nghiên cứu rộng rãi trước đó, và quan trọng hơn, chúng cho thấy có liên kết với các hoạt tính sinh học của thực vật chủ 5 . Vì cũng là vi sinh vật nên các vi khuẩn thuộc nhóm Streptomyces cũng cho thấy đặc tính này. Ở các vi khuẩn Streptomyces , hoạt tính sinh học nói chung và hoạt tính kháng khuẩn nói riêng được quy định bởi các hợp chất thứ cấp (secondary metabolites) 6 . Ở Streptomyces , các hợp chất thứ cấp được sinh tổng hợp bởi các nhóm gen (gene cluster) tại các locus xác định trên bộ gen 7 . Vì vậy, việc xác định các gene cluster chịu trách nhiệm sinh tổng hợp các hợp chất thứ cấp có hoạt tính sinh học ở Streptomyces không những cần thiết cho các nghiên cứu cơ bản nhằm khám phá con đường sinh tổng hợp các hợp chất thứ cấp mà còn cho các nghiên cứu ứng dụng để sản xuất hợp chất thứ cấp có hoạt tính sinh học.
Nhằm tạo tiền đề cho các nghiên cứu cơ bản và ứng dụng tiếp theo trên chủng Streptomyces có hoạt tính sinh học, trước tiên cần phải truyền được DNA ngoại lai vào tế bào chủng Streptomyces mục tiêu vì đây là cơ sở để thực hiện các thí nghiệm nghiên cứu chức năng gen bao gồm gây đột biến gen và bổ trợ chủng đột biến bằng bản sao gen bị đột biến. Ở Streptomyces , có các phương pháp thường được sử dụng để truyền DNA ngoại lai vào bên trong tế bào như: hoá biến nạp với polyethylene glycol, điện biến nạp, và tiếp hợp vi khuẩn. Trong số các phương pháp này, tiếp hợp thường được sử dụng để truyền DNA ngoại lai trong các thí nghiệm gây đột biến gen bằng kỹ thuật tái tổ hợp tương đồng 8 , biểu hiện gen 9 , chỉnh sửa bộ gen bằng công nghệ CRISPR/Cas9 10 ở nhóm vi khuẩn Streptomyces . Phương pháp truyền DNA ngoại lai bằng tiếp hợp vi khuẩn cho thấy các ưu điểm về thao tác, thời gian tiến hành, chi phí so với các phương pháp truyền DNA ngoại lai còn lại như hoá biến nạp, điện biến nạp.
Để đáp ứng với xu hướng tìm kiếm kháng sinh mới hiện nay, trong nghiên cứu này, chúng tôi đã phân lập được một chủng Streptomyces nội sinh từ cây Hoa giấy và chủng Streptomyces này đã cho thấy hoạt tính kháng khuẩn đáng chú ý trên các chủng vi khuẩn gây bệnh. Chúng tôi cũng đã thiết lập các điều kiện để chuyển DNA ngoại lai vào tế bào của chủng Streptomyces nhằm tạo tiền đề cho các nghiên cứu về hoạt tính kháng khuẩn về sau trên chủng Streptomyces này.
VẬT LIỆU VÀ PHƯƠNG PHÁP
Vi sinh vật, plasmid và primer
Các vi khuẩn gây bệnh bao gồm Enterococcus faecium , Staphylococcus aureus , Klebsiella pneumoniae , Acinetobacter baumannii , Pseudomonas aeruginosa , Enterobacter cloacae , Escherichia coli là các chủng vi khuẩn lâm sàng do Trung tâm Nghiên cứu và Ứng dụng Sinh học cung cấp. ET12567/pUZ8002 là quà tặng của Tiến sĩ Nguyễn Kim Nữ Thảo (Bộ sưu tập Chủng vi sinh vật Việt Nam-VTCC). Chủng do PGS.TS. Đặng Thị Phương Thảo (Trường ĐH Khoa học Tự nhiên, ĐHQG-HCM) cung cấp. Chủng được cung cấp từ Viện Pasteur Thành phố Hồ Chí Minh. Plasmid pOSV206 do Tiến sĩ Jean-Luc Pernodet (Viện I2BC, Pháp) cung cấp. Cặp primer 27F-1495R để khuếch đại gen 16S rRNA của vi khuẩn và cặp primer i.aacF (CGGATCTCGGCCCAGTTGAC) và i.aacR (CGTTCGATCCTGCCACGTCG) để khuếch đại gen aac được mua từ công ty PhuSa Biochem (Việt Nam).
Môi trường nuôi cấy, kháng sinh và hoá chất
Môi trường Nutrient broth được sử dụng để nuôi cấy vi khuẩn gây bệnh. Môi trường Mueller-Hinton agar (MHA) được sử dụng để kiểm tra hoạt tính kháng khuẩn. Môi trường Luria-Bertani (LB) được sử dụng để nuôi cấy , và ET12567/pUZ8002. Soya Flour Mannitol (SFM) được sử dụng để phân lập chủng vi khuẩn nội sinh. FM3 là môi trường nuôi cấy sinh hoạt tính kháng khuẩn. Streptomyces sp. SS200 được nuôi cấy trong YEME để thu nhận khuẩn ty cho thí nghiệm tiếp hợp và tách chiết DNA bộ gen. SFM, Heskey-Tresner (HT), Trypticase Soy Agar (TSA) và 2CMC là các môi trường tiếp hợp. Các môi trường nuôi cấy ISP (International Streptomyces Project) cùng với casein agar, MacConkey và Simmons citrate agar được sử dụng để xác định hình thái của SS200. Axit nalidixic, apramycin và kanamycin được mua từ GoldBio (Mỹ). Hoá chất cho PCR và các hoá chất khác được mua từ công ty TBR (Việt Nam).
Phân lập chủng vi khuẩn nội sinh
Lá của cây Hoa giấy ( Bougainvillea glabrai ) được thu hái tại thành phố Vũng Tàu, Việt Nam và vận chuyển đến phòng thí nghiệm trong vòng 2-3 giờ. Chủng vi khuẩn nội sinh được phân lập theo quy trình của Nguyễn Đăng Quân và cộng sự 7 . Theo đó, mẫu lá được rửa bằng nước để loại bỏ tạp chất trên bề mặt và sau đó trải qua nhiều lần rửa với 0,1% Tween 20, 5% NaOCl, 5% Na 2 S 2 O 3 , 70% ethanol, nước cất khử trùng và dung dịch NaHCO 3 10% để khử trùng bề mặt của mẫu lá. Mẫu lá được nghiền trong NaCl 0,9% vô trùng, sau đó dịch chiết thực vật được trải trên đĩa thạch SFM có bổ sung axit nalidixic, cycloheximide và nystatin ở nồng độ 50 mg/ml cho mỗi loại kháng sinh. Các đĩa SFM được ủ ở 28 o C cho đến khi các khuẩn lạc Streptomyces điển hình xuất hiện trên bề mặt thạch SFM. Các khuẩn lạc được làm thuần bằng cách cấy ria nhiều lần trên môi trường SFM. Các khuẩn lạc thuần được giữ trong 30% glycerol ở -20 o C để bảo quản cho các thí nghiệm tiếp theo.
Khảo sát hoạt tính kháng khuẩn của chủng vi khuẩn đã phân lập
Các chủng vi khuẩn gây bệnh được nuôi cấy lắc trong môi trường Nutrient broth ở 37 o C qua đêm. Mỗi dịch vi khuẩn được pha loãng với dung dịch 0,9% NaCl vô trùng đến nồng độ cuối là 1x10 8 CFU/ml. 100 ml của mỗi dịch vi khuẩn được trải trên các đĩa MHA. Sau đó, các lỗ được đục một cách vô trùng bằng các đầu tip để tạo thành các giếng trên bề mặt các đĩa MHA. Chủng phân lập, được đặt tên là SS200, được nuôi cấy lắc trong môi trường FM3 ở 28 o C trong 5 ngày. Sau đó, dịch môi trường nuôi cấy SS200 được sử dụng để kiểm tra hoạt tính kháng khuẩn bằng phương pháp khuếch tán giếng thạch 11 . Dịch nuôi cấy SS200 được cho vào các giếng trên các đĩa MHA ở trên, và các đĩa này được ủ ở 37 o C trong 16-18 giờ. Hoạt tính kháng khuẩn của SS200 đối với các vi khuẩn gây bệnh được ghi nhận bằng cách đo đường kính các vòng vô khuẩn xung quanh các giếng. Dịch môi trường FM3 và kháng sinh apramycin (1 mg/ml) được sử dụng làm đối chứng âm và đối chứng dương trong thí nghiệm này.
Định danh chủng SS200
Để xác định đặc điểm hình thái, chủng vi khuẩn SS200 được nuôi cấy trên nhiều loại môi trường ISP khác nhau 12 . Các đĩa môi trường đã cấy với SS200 được ủ ở 28 o C trong 7 ngày. Sự phát triển, hình thái khuẩn lạc và màu sắc bào tử của SS200 được ghi nhận.
Để định danh phân tử, chủng SS200 được nuôi cấy lắc trong môi trường YEME ở 28 o C trong 3 ngày. Khuẩn ty của SS200 được thu nhận và rửa sạch với nước cất khử trùng để loại bỏ các thành phần môi trường nuôi cấy. DNA bộ gen được tách chiết từ khuẩn ty SS200 bằng phương pháp phenol/chloroform và DNA này được khuếch đại bằng PCR với cặp primer 27F-1495R là cặp primer khuếch đại gen 16S rRNA của vi khuẩn 13 . Sản phẩm PCR thu được đã được tinh chế qua gel agarose và giải trình tự bằng công nghệ Sanger. Trình tự nucleotide của sản phẩm PCR được chỉnh sửa bằng phần mềm BioEdit và tải lên ngân hàng dữ liệu trên GenBank. Ngoài ra, trình tự gen 16S rRNA của SS200 được phân tích bằng công cụ 16S-based ID của cơ sở dữ liệu EZBioCloud tại địa chỉ www.ezbiocloud.net để định danh phân tử chủng SS200 14 .
Thí nghiệm tiếp hợp cơ bản
Sự tiếp hợp giữa E. coli và Streptomyces sp. SS200 được thực hiện theo công trình của Kiesser và cộng sự 15 với các thay đổi nhỏ. Khuẩn ty của chủng Streptomyces được sử dụng trong thí nghiệm tiếp hợp thay cho bào tử. Thí nghiệm tiếp hợp được thực hiện với các điều kiện cơ bản bao gồm ET12567/pUZ8002 là chủng cho DNA, SFM làm môi trường tiếp hợp, MgCl 2 ở nồng độ 30 mM và thời gian ủ trước khi chọn lọc kháng sinh là 24 giờ. Streptomyces sp. SS200 được nuôi cấy lắc trong môi trường YEME ở 28 o C trong 2 ngày. Sau đó, 100 mg khuẩn ty được trộn với 1x10 9 CFU/ml E. coli . Hỗn dịch giữa hai loại vi khuẩn này được trải trên các đĩa thạch SFM có chứa 30 mM MgCl 2 và thời gian ủ là 24 giờ trước khi áp dụng chọn lọc với kháng sinh là axit nalidixic và apramycin. Tần số tiếp hợp được tính bằng cách chia số lượng exconjugant cho số lượng tế bào E. coli cho DNA 16 . Các điều kiện tiếp hợp cơ bản sẽ được sử dụng để sàng lọc các điều kiện tiếp hợp cho tần số tiếp hợp cao nhất.
Sàng lọc các điều kiện tối ưu cho tiếp hợp
Từ các điều kiện tiếp hợp cơ bản, các điều kiện tiếp hợp như chủng E. coli cho DNA, các cation 2+ , môi trường tiếp hợp, thời gian ủ trước khi chọn lọc kháng sinh được sàng lọc để thu được điều kiện tiếp hợp tối ưu. Ngoài ET12567/pUZ8002, chủng E. coli cho DNA khác là được sử dụng trong tiếp hợp với Streptomyces sp. SS200. Ảnh hưởng của cation 2+ được khảo sát với hai loại muối là MgCl 2 và CaCl 2 . Nồng độ muối khảo sát nằm trong khoảng từ 0-50 mM cho mỗi loại muối. Bốn môi trường tiếp hợp bao gồm SFM, HT, TSA, 2CMC và các thời gian ủ trước khi chọn lọc kháng sinh là 0, 6, 12, 18 và 24 giờ được sàng lọc để chọn ra môi trường tiếp hợp và thời gian ủ cho hiệu suất tiếp hợp cao nhất. Các thí nghiệm tối ưu hóa được thực hiện theo kiểu đơn yếu tố. Tất cả các thí nghiệm được thực hiện ba lần lặp lại để lấy giá trị trung bình.
Xác nhận các thể tiếp hợp (exconjugant) bằng PCR
Các thể tiếp hợp từ sự tiếp hợp giữa E. coli và Streptomyces sp. SS200 được nuôi cấy trong môi trường YEME và khuẩn ty của chúng được sử dụng để tách chiết DNA bộ gen. DNA bộ gen được sử dụng làm khuôn mẫu trong hai phản ứng PCR với cặp mồi 27F-1495R để khuếch đại gen 16S rRNA của Streptomyces sp. SS200 và cặp mồi i.aacF-1.aacR để khuếch đại đoạn DNA 500 bp của gen aac trên plasmid pOSV206. Các sản phẩm PCR thu được được kiểm tra bằng điện di trên gel agarose và kích thước của các sản phẩm PCR được so sánh với thang DNA 100 bp và thang DNA lHindIII.
KẾT QUẢ
Phân lập chủng vi khuẩn SS200 từ Bougainvillea glabra
Một chủng vi khuẩn nội sinh đã được phân lập từ lá của cây Hoa giấy Bougainvillea glabra thu nhận từ thành phố Vũng Tàu. Ở bước phân lập, chủng vi khuẩn này đề kháng với axit nalidixic, cycloheximide và nystatin, đây là các kháng sinh ức chế các vi khuẩn Gram âm và nấm. Ngoài ra, chủng vi khuẩn này cho các khuẩn lạc được cấu tạo bởi các khuẩn ty cơ chất và khuẩn ty khí sinh chứa các bào tử màu xám trên môi trường thạch SFM (Hình 1). Những đặc điểm kể trên là đặc trưng cho vi khuẩn Streptomyces trên môi trường nuôi cấy rắn. Với kết quả ban đầu này, chủng vi khuẩn đã phân lập được đặt tên là SS200 và được sử dụng để khảo sát khả năng kháng khuẩn trên các chủng vi khuẩn gây bệnh kháng kháng sinh ở thí nghiệm tiếp theo.

Hoạt tính kháng khuẩn của chủng vi khuẩn SS200 đã phân lập
Các vi khuẩn gây bệnh kháng kháng sinh trong thí nghiệm này thuộc các nhóm khác nhau dựa trên đặc tính nhuộm Gram và hình dạng vi khuẩn. Hơn nữa, hầu hết chúng thuộc về nhóm ESKAPE, bao gồm Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, Enterobacter cloacae . Kết quả khảo sát cho thấy chủng vi khuẩn SS200 thể hiện phổ kháng khuẩn rộng trên tất cả các vi khuẩn gây bệnh (Hình 2).
Figure 2 . Hoạt tính kháng khuẩn của SS200 đối với các chủng vi khuẩn gây bệnh. FM3: dịch môi trường FM3; SS200/FM3-1,-2,-3: Dịch môi trường FM3 có nuôi cấy SS200; Apr: kháng sinh chứng dương apramycin
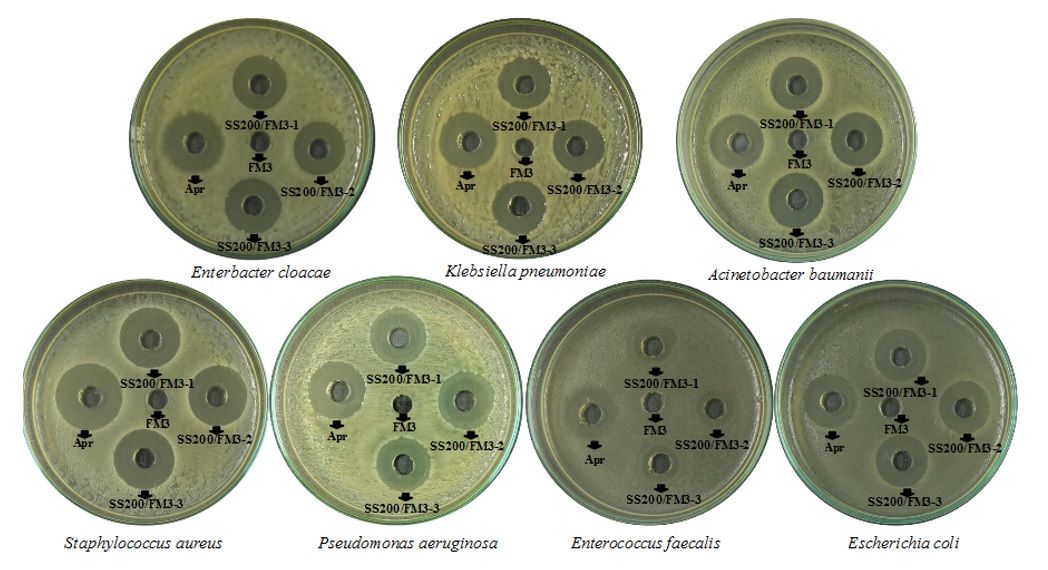
Vòng vô khuẩn đối với các vi khuẩn gây bệnh nằm trong khoảng từ 15 mm đến 25 mm. Để so sánh, chứng dương apramycin ở nồng độ 1 mg/ml cho vòng vô khuẩn nằm trong khoảng từ 21 mm đến 26 mm (Bảng 1).
Đặc tính hình thái và phân tử của chủng vi khuẩn SS200
Hình 3 cho thấy hình thái của chủng vi khuẩn SS200 trên các môi trường nuôi cấy, trong đó có các môi trường nuôi cấy đặc trưng cho nhóm vi khuẩn Streptomyces (các môi trường ISP). SS200 phát triển trên tất cả các môi trường nuôi cấy ngoại trừ môi trường MacConkey, là môi trường nuôi cấy chọn lọc cho vi khuẩn Gram âm. Trên các môi trường nuôi cấy, SS200 cho thấy sự phát triển trung bình cho đến mạnh. Hình thái khuẩn lạc rất đa dạng bao gồm hình tròn và các hình dạng bất thường khác. Bề mặt của khuẩn lạc có thể là bằng phẳng, thô ráp, lồi lõm hoặc nổi lên ở tâm. Màu sắc của bào tử thay đổi tùy theo môi trường nuôi cấy, bao gồm các màu xám, trắng hoặc vàng lục nhạt.

DNA bộ gen được chiết xuất từ khuẩn ty của SS200 sau đó được nhân bản bằng PCR với cặp primer 27F-1495R để thu nhận sản phẩm gen 16S rRNA. Sản phẩm PCR thu được được giải trình tự bằng công nghệ Sanger, trình tự nucleotide được hiệu chỉnh để thu nhận đoạn trình tự có kích thước 1.376 nucleotide. Trình tự nucleotide này được gửi vào ngân hàng dữ liệu nucleotide trên GenBank và được cấp số truy cập OM855561. Ngoài ra, trình tự nucleotide này được sử dụng để định danh phân tử chủng SS200 bằng công cụ 16S-based ID của cơ sở dữ liệu về phân loại EZBioCloud. Kết quả cho thấy trình tự 16S rRNA của chủng SS200 có độ tương đồng hơn 99% so với trình tự 16S rRNA của các chủng Streptomyces trong cơ sở dữ liệu (Hình 4).

Như vậy, đặc điểm hình thái và phân tử xác định SS200 là một chủng vi khuẩn thuộc nhóm Streptomyces . Do đó, chủng vi khuẩn này được đặt tên là Streptomyces sp. SS200.
Thí nghiệm tiếp hợp cơ bản giữa E. coli và Streptomyces sp. SS200
Trong thí nghiệm tiếp hợp cơ bản để truyền DNA vào Streptomyces sp. SS200, các điều kiện tiếp hợp cơ bản bao gồm dạng tế bào nhận là khuẩn ty SS200, ET12567/pUZ8002 là chủng E. coli cho DNA, MgCl 2 ở nồng độ 30 mM là dạng muối vô cơ cần thiết cho sự tiếp hợp vi khuẩn, SFM là môi trường tiếp hợp và 24 giờ là thời gian ủ trước khi chọn lọc với kháng sinh. Kết quả của thí nghiệm tiếp hợp cơ bản trong Hình 5 cho thấy các thể tiếp hợp điển hình của Streptomyces sp. SS200 xuất hiện trên mặt môi trường SFM chứa axit nalidixic và apramycin. Số lượng các thể tiếp hợp trung bình trong thí nghiệm tiếp hợp cơ bản là 64,3±4,1, dựa trên đó tần số tiếp hợp được tính ra là 2,04x10 -8 .
Figure 5 . Các thể tiếp hợp từ thí nghiệm tiếp hợp giữa chủng ET12567/pUZ8002 mang pOSV206 và Streptomyces sp. SS200
Ảnh hưởng của các điều kiện tiếp hợp đến sự tiếp hợp ở Streptomyces sp. SS200
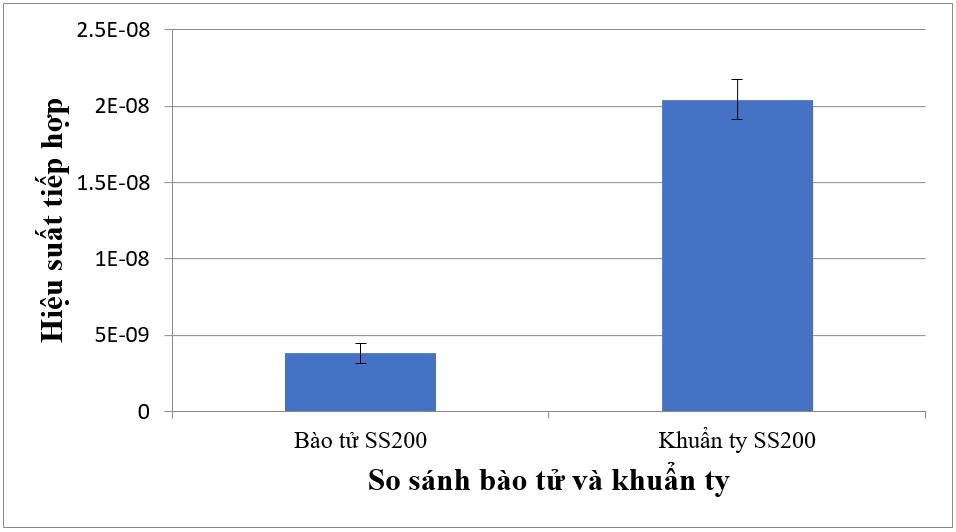
Ngoài dạng khuẩn ty Streptomyces sp. SS200, bào tử của vi khuẩn này cũng được khảo sát như dạng tế bào nhận DNA. Trong số hai dạng tế bào nhận DNA của Streptomyces sp. SS200, dạng khuẩn ty cho tần số tiếp hợp là 2,04x10 -8 , tần số tiếp hợp này cao hơn tần số tiếp hợp của dạng bào tử Streptomyces sp. SS200 là 3,8x10 -9 (Hình 6).
Figure 6 . So sánh hiệu quả dạng tế bào nhận của Streptomyces sp. SS200 đối với sự tiếp hợp
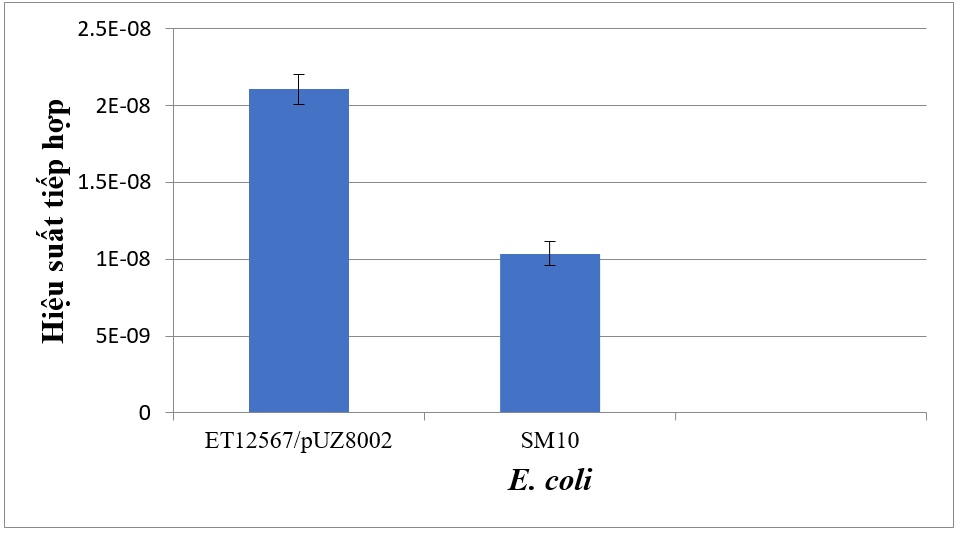
Ngoài chủng E. coli ET12567/pUZ8002 trong thí nghiệm tiếp hợp cơ bản, một chủng E. coli cho DNA khác là cũng được khảo sát khả năng truyền DNA vào chủng Streptomyces sp. SS200. Kết quả trong Hình 6 cho thấy chủng vi khuẩn ET12567/pUZ8002 là chủng vi khuẩn E. coli cho tần số tiếp hợp cao nhất là 2,1x10 -8 trong khi cho tần số tiếp hợp là 1,03x10 -8 (Hình 7).
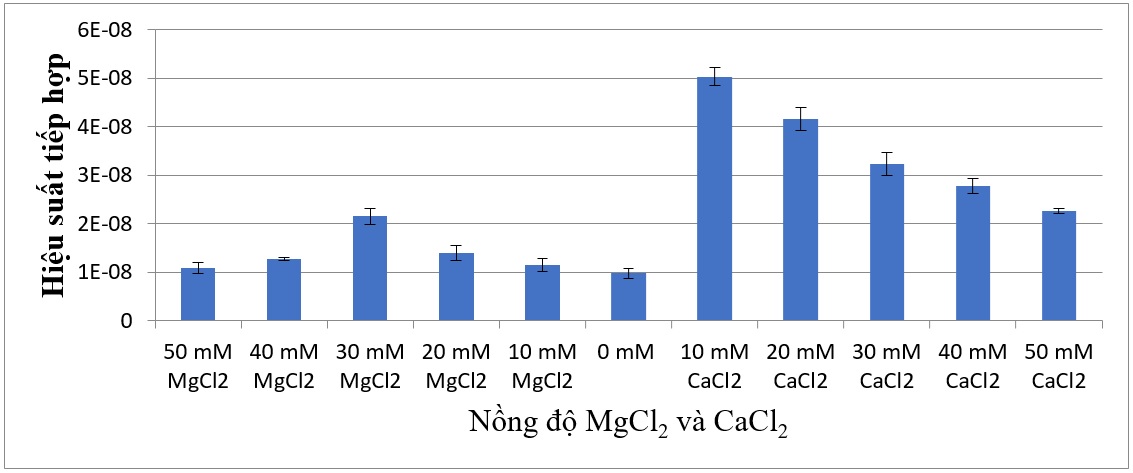
Hai loại muối vô cơ thường được sử dụng trong các thí nghiệm tiếp hợp ở vi khuẩn là MgCl 2 và CaCl 2 cũng được khảo sát trong sự tiếp hợp giữa E. coli và Streptomyces sp. SS200. Mỗi muối được khảo sát riêng lẻ trong dãy nồng độ từ 0 đến 50 mM với mức tăng là 10 mM trong môi trường tiếp hợp. Ở 0 mM MgCl 2 , sự tiếp hợp giữa E. coli và Streptomyces sp. SS200 vẫn xảy ra nhưng ở tần suất tiếp hợp thấp nhất là 9,11x10 -9 . Tần số tiếp hợp tăng khi tăng nồng độ MgCl 2 trong môi trường tiếp hợp và đạt đến tần số 2,14x10 -8 ở nồng độ 30 mM MgCl 2 . Các nồng độ MgCl 2 cao hơn 30 mM gây giảm tần số tiếp hợp, cụ thể là 1,26x10 -8 ở nồng độ 40 mM MgCl 2 và 1,07x10 -8 ở nồng độ 50 mM MgCl 2 (Hình 8).

Đối với muối CaCl 2 , tần số tiếp hợp cao nhất là 5,02x10 -8 được tìm thấy ở nồng độ 10 mM CaCl 2 và tần số tiếp hợp giảm khi nồng độ CaCl 2 tăng lên (Hình 9).

Khi so sánh ảnh hưởng của hai muối vô cơ MgCl 2 và CaCl 2 trên tần số tiếp hợp giữa E. coli và Streptomyces sp. SS200 thì các nồng độ CaCl 2 cho tần số tiếp hợp cao hơn so với các nồng độ MgCl 2 (Hình 10).
Figure 10 . So sánh hiệu quả các nồng độ muối MgCl 2 và CaCl 2 đối với sự tiếp hợp
Bốn môi trường tiếp hợp bao gồm SFM, TSA, HT và 2CMC cũng được khảo sát để tìm ra môi trường cho tần số tiếp hợp cao nhất giữa E. coli và Streptomyces sp. SS200. Trong số bốn môi trường này, các thể tiếp hợp chỉ xuất hiện trên môi trường SFM với tần số tiếp hợp là 1,97x10 -8 trong khi ba môi trường tiếp hợp còn lại không có sự xuất hiện bất kỳ thể tiếp hợp nào (Hình 11).

Cuối cùng, các thời gian ủ bao gồm 0, 6, 12, 18 và 24 giờ trước khi chọn lọc với kháng sinh cũng được khảo sát nhằm tìm ra thời gian ủ cho tần số tiếp hợp cao nhất giữa E. coli và Streptomyces sp. SS200. Kết quả khảo sát cho thấy tần số tiếp hợp tăng lên với thời gian ủ dài hơn, tần suất tiếp hợp cao nhất là 1,97x10 -8 với thời gian ủ là 24 giờ sau tiếp hợp. Không có bất kỳ thể tiếp hợp nào trên môi trường tiếp hợp với thời gian ủ là 0 giờ (Hình 12).

Như vậy, dạng khuẩn ty của Streptomyces sp. SS200, chủng vi khuẩn E. coli ET12567/pUZ8002, muối CaCl 2 ở nồng độ 10 mM, môi trường tiếp hợp là SFM, thời gian ủ 24 giờ trước khi chọn lọc với kháng sinh là các điều kiện được chọn cho các thí nghiệm tiếp hợp trong nghiên cứu chức năng gen về sau trên chủng của Streptomyces sp. SS200.
Xác nhận các thể tiếp hợp giữa E. coli và Streptomyces sp. SS200 bằng PCR
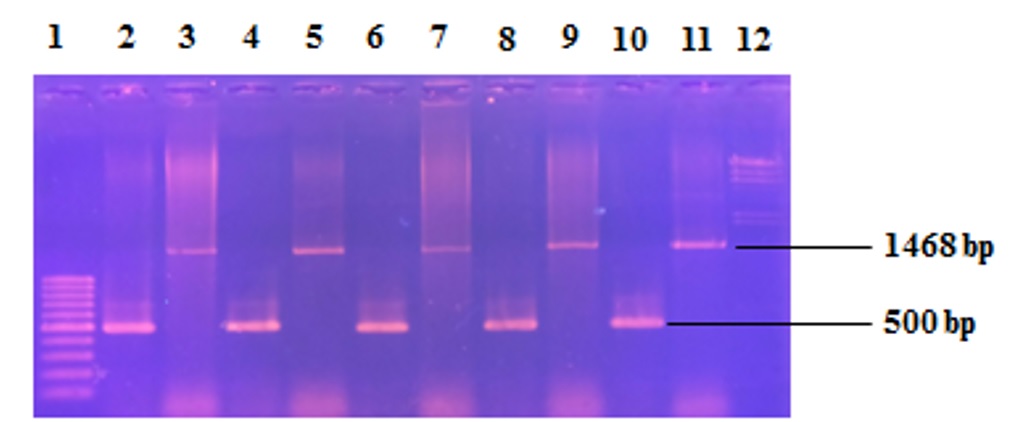
Năm thể tiếp hợp giữa E. coli và Streptomyces sp. SS200 được chọn ngẫu nhiên để xác nhận bằng PCR với hai cặp primer, một cặp primer cho gen 16S rRNA trong bộ gen của Streptomyces sp. SS200 và một cặp primer để khuếch đại gen aac trên plasmid pOSV206. Hình 13 cho thấy kết quả điện di trên gel agarose các sản phẩm PCR từ năm thể tiếp hợp đã khảo sát. Mỗi thể tiếp hợp cho hai sản phẩm PCR với kích thước là 1.468 bp (đối với gen 16S rRNA) và 500 bp (đối với gen aac ) khi so sánh với thang DNA 100 bp và thang DNA lHindIII.
Figure 13 . Xác nhận các thể tiếp hợp giữa E. coli và Streptomyces sp. SS200 bằng PCR. 1: thang DNA 100 bp; 2,4,6,8,10: sản phẩm PCR với cặp primer 27F-1942R từ năm thể tiếp hợp; 3,5,7,9,11: sản phẩm PCR với cặp primer i.aacF-i.aacR từ năm thể tiếp hợp; 12: thang DNA lHindIII
THẢO LUẬN
Trong các nghiên cứu khảo sát hoạt tính sinh học khác, Bougainvillea glabra cho thấy có các hoạt tính sinh học đa dạng như chống tiểu đường, gây độc tế bào, kháng viêm, kháng oxi hoá và đặc biệt là kháng vi sinh vật 17 . Gần đây, một giả thuyết về sự trao đổi vật liệu di truyền giữa thực vật và các sinh vật nội sinh của chúng đã được đề xuất để giải thích cho hiện tượng thực vật và vi sinh vật nội sinh có cùng các hoạt tính sinh học thông qua việc cùng sản xuất các hợp chất thứ cấp tương ứng 18 . Vì vậy, có khả năng các vi sinh vật nội sinh trong Bougainvillea glabra cũng có thể sản sinh các hoạt tính sinh học tương tự như thực vật chủ và đây cũng là lý do để phân lập và khảo sát hoạt tính kháng khuẩn các chủng vi sinh vật nội sinh trong Bougainvillea glabra . Trong nghiên cứu này, một chủng vi sinh vật nội sinh, được đặt tên là SS200, đã được phân lập từ cây Hoa giấy ( Bougainvillea glabra ) và chủng vi sinh vật này cho thấy hoạt tính kháng khuẩn phổ rộng đối với các vi khuẩn gây bệnh thuộc nhóm ESKAPE.
Sau khi phân lập từ Bougainvillea glabra , chủng vi sinh vật SS200 được cho là thuộc nhóm vi khuẩn Gram dương vì chủng này có khả năng kháng lại các kháng sinh cycloheximide và nystatin, là các kháng sinh ức chế sự phát triển của nấm. SS200 cũng có khả năng kháng axit nalidixic, là kháng sinh ức chế vi khuẩn Gram âm. Ngoài ra, khuẩn lạc của SS200 có cấu tạo bởi khuẩn ty cơ chất ăn sâu xuống mặt môi trường SFM và khuẩn ty khí sinh mang chuỗi bào tử. Đây là những đặc tính kiểu hình đặc trưng cho nhóm vi khuẩn Streptomyces . Trên các môi trường ISP, SS200 tiếp tục cho thấy các đặc tính khuẩn lạc đặc trưng cho nhóm vi khuẩn Streptomyces . Tiếp theo, chúng tôi thực hiện việc định danh phân tử chủng SS200 dựa trên gen 16S rRNA. Trong bước định danh này chúng tôi sử dụng cặp primer 27F-1495R để nhân bản gen 16S rRNA của SS200. 27F-1495R là cặp primer được sử dụng để nhân bản gen 16S rRNA của giới vi khuẩn cho mục tiêu định danh phân tử trong nhiều nghiên cứu. Trình tự nucleotide gen 16S rRNA của SS200 cho thấy có sự tương đồng cao với gen 16S rRNA của các chủng Streptomyces trong cơ sở dữ liệu như S. rochei , S. enissocaesilis , S. plicatus , S. geysiriensis , v.v. Để xác định chính xác mức độ loài thuộc giống Streptomyces đối với chủng vi khuẩn SS200 thì cần có nghiên cứu phân loại hệ thống (polyphasic taxonomy) trên nhiều đặc tính khác nhau như nuôi cấy, sinh lý và sinh hóa, tành phân cấu tạo thành tế bào và màng tế bào, hình thái tế bào dưới kính hiển vi điện tử, giải trình tự và so sánh bộ gen với dữ liệu bộ gen của các chủng Streptomyces . Kết quả từ nghiên cứu phân loại hệ thống này sẽ giúp phân loại một chủng Streptomyces đến mức loài 19 . Do nghiên cứu này không thực hiện việc phân loại hệ thống chủng SS200 và dựa trên các kết quả về hình thái và phân tử thu được trong nghiên cứu này, chủng vi khuẩn SS200 được đặt tên là Streptomyces sp. SS200.
Phổ kháng khuẩn rộng của Streptomyces sp. SS200 tạo tiền đề cho các nghiên cứu tiếp theo nhằm phân lập hợp chất thứ cấp có hoạt tính kháng khuẩn cũng như nghiên cứu con đường sinh tổng hợp hợp chất thứ cấp này. Vì vậy ở bước kế tiếp, chúng tôi thiết lập các điều kiện để truyền DNA vào chủng Streptomyces sp. SS200 bằng phương pháp tiếp hợp. Phương pháp tiếp hợp giữa E. coli và Streptomyces đã được Mazodier và cộng sự thiết lập vào năm 1989 20 và kể từ đó, các điều kiện khác nhau đã được xác định có ảnh hưởng đến hiệu suất tiếp hợp như: chủng E. coli cho DNA, dạng tế bào Streptomyces nhận DNA, loại muối vô cơ và nồng độ của chúng, môi trường tiếp hợp, thời gian ủ trước khi chọn lọc với kháng sinh. Các điều kiện này thường được khảo sát để tìm ra thông số tối ưu cho từng thí nghiệm tiếp hợp cụ thể ở Streptomyces . Vì vậy, trong nghiên cứu này, các điều kiện này cũng được khảo sát để tìm ra các thông số tối ưu cho thí nghiệm tiếp hợp ở chủng Streptomyces sp. SS200. Đối với điều kiện về chủng E. coli cho DNA, chủng ET12567/pUZ8002 cho thấy hiệu quả tiếp hợp tốt hơn chủng . Chủng ET12567/pUZ8002 là chủng bị khiếm khuyết trong khả năng methyl hoá DNA trong khi lại là chủng có khả năng methyl hoá DNA hoàn toàn. Tình trạng methyl hoá của DNA cho có tác động quan trọng đối với sự tiếp hợp giữa E. coli và Streptomyces . Nhiều chủng Streptomyces có hệ thống cắt giới hạn DNA cho bằng enzyme phụ thuộc vào tình trạng methyl của DNA cho 21 . Những hệ thống cắt giới hạn như vậy làm giảm hiệu suất truyền DNA vào tế bào bằng tất cả các phương pháp như hoá biến nạp, điện biến nạp và tiếp hợp. Trong những chủng Streptomyces như vậy, chủng E. coli cho DNA thường được sử dụng là ET12567/pUZ8002. Trong những chủng Streptomyces không có hệ thống cắt giới hạn DNA cho bằng enzyme phụ thuộc vào tình trạng methyl của DNA cho, có thể thay thế cho ET12567/pUZ8002 làm chủng cho DNA trong các thí nghiệm tiếp hợp.
Cả hai dạng tế bào Streptomyces nhận DNA là bào tử và khuẩn ty đều có thể được sử dụng trong thí nghiệm tiếp hợp với E. coli . Hiệu suất tiếp hợp của hai dạng tế bào này thay đổi tuỳ theo từng chủng Streptomyces cụ thể. Dạng bào tử của Streptomyces được thấy cho hiệu suất tiếp hợp cao hơn dạng khuẩn ty ở Streptomyces rimosus M527 22 . Ngược lại, dạng khuẩn ty lại cho hiệu suất tiếp hợp cao hơn dạng bào tử ở Streptomyces kanamyceticus ATCC 12853 23 , kết quả trong nghiên cứu này cũng cho thấy dạng khuẩn ty của Streptomyces sp. SS200 cho hiệu suất tiếp hợp hiệu quả hơn dạng bào tử. Ngoài ra, ở những chủng Streptomyces không tạo được bào tử thì khuẩn ty là dạng tế bào nhận duy nhất trong thí nghiệm tiếp hợp 24 . Ngoài hai điều kiện chủng cho và chủng nhận DNA, các điều kiện tiếp hợp còn lại cũng được khảo sát để tìm ra điều kiện cho hiệu suất tiếp hợp cao nhất. Trong số hai loại muối vô cơ thì CaCl 2 cho hiệu suất cao hơn MgCl 2 trên tất cả các nồng độ đi từ 10 mM đến 50 mM. Ở nồng độ 10 mM, CaCl 2 có hiệu suất tiếp hợp cao nhất là 5,02x10 -8 . CaCl 2 cũng là muối vô cơ cho hiệu suất tiếp hợp cao hơn MgCl 2 ở chủng Streptomyces neptrosis SD-07, Streptomyces coelicolor , Streptomyces lavendulae và Streptomyces venezuelae 25 . Tương tự, hai điều kiện tiếp hợp còn lại là môi trường nuôi cấy và thời gian ủ trước khi chọn lọc với kháng sinh cũng cho thấy ảnh hưởng đối với hiệu suất tiếp hợp ở chủng Streptomyces sp. SS200. Cụ thể, môi trường SFM và thời gian ủ 24 giờ trước khi chọn lọc với kháng sinh là những điều kiện tiếp hợp cho các hiệu suất tiếp hợp giữa E. coli và Streptomyces sp. SS200 cao nhất.
KẾT LUẬN
Streptomyces sp. SS200 được phân lập từ cây Hoa giấy ( Bougainvillea glabra ) thu hái ở thành phố Vũng Tàu, Việt Nam, và chủng vi khuẩn này cho thấy hoạt tính kháng khuẩn đối với một số vi khuẩn gây bệnh thuộc nhóm ESKAPE. Tiếp theo, các điều kiện để chuyển DNA vào chủng Streptomyces sp. SS200 bằng phương pháp tiếp hợp được khảo sát để thu nhận thông số tối ưu cho từng điều kiện tiếp hợp. Các thông số tối ưu đã khảo sát bao gồm: dạng tế bào nhận DNA của Streptomyces sp. SS200 là khuẩn ty, ET12567/pUZ8002 là chủng E. coli cho DNA, CaCl 2 ở nồng độ 10 mM, SFM là môi trường tiếp hợp và thời gian ủ là 24 giờ trước khi chọn lọc với kháng sinh. Những điều kiện tiếp hợp này sẽ được sử dụng trong các thí nghiệm tiếp theo nhằm xác định các gen liên quan đến con đường sinh tổng hợp của các chất chuyển hóa thứ cấp có hoạt tính kháng khuẩn ở Streptomyces sp. SS200.
CẢM TẠ
Các tác giả chân thành cám ơn Trung tâm Nghiên cứu và Ứng dụng Sinh học đã cung cấp các vật liệu, hoá chất và thiết bị để thực hiện nghiên cứu. Các tác giả cũng chân thành cám ơn Quỹ phát triển Khoa học và Công nghệ NTTU đã tài trợ một phần kinh phí cho nghiên cứu này trong đề tài mã số 2022.01.09/HĐ-KHCN.
XUNG ĐỘT LỢI ÍCH
Các tác giả đồng ý, không có bất kỳ xung đột lợi ích nào liên quan đến các kết quả đã công bố.
ĐÓNG GÓP CỦA CÁC TÁC GIẢ
Nguyễn Thị Ngọc Quyên lên kế hoạch, thực hiện các thí nghiệm. Nguyễn Trung Hiếu đánh giá, xử lý các dữ liệu. Nguyễn Hoàng Chương định hướng, chỉnh sửa và viết bản thảo.
References
- . . ;:. Google Scholar
- Tagliabue A, Rappuoli R. Changing Priorities in Vaccinology: Antibiotic Resistance Moving to the Top. Front Immunol. 2018;9:1068. . ;:. PubMed Google Scholar
- Duval RE, Grare M, Demoré B. Fight Against Antimicrobial Resistance: We Always Need New Antibacterials but for Right Bacteria. Molecules. 2019;24:3152. . ;:. PubMed Google Scholar
- Hopwood DA. Streptomyces in Nature and Medicine. The Antibiotic Makers. New York, NY: Oxford University Press Inc. 2007. . ;:. Google Scholar
- Dhayanithy G, Subban K & Chelliah J. Diversity and biological activities of endophytic fungi associated with Catharanthus roseus. BMC Microbiol. 2019;19:22. . ;:. PubMed Google Scholar
- Lacey HJ, Rutledge PJ. Recently Discovered Secondary Metabolites from Streptomyces Species. Molecules. 2022;27:887. . ;:. PubMed Google Scholar
- Nguyen QD, Truong PM, Vo TNT, Chu TDX, Nguyen CH. Draft genome sequence data of Streptomyces sp. SS1-1, an endophytic strain showing cytotoxicity against the human lung cancer A549 cell line. Data Brief. 2020;30:105497. . ;:. PubMed Google Scholar
- Nguyen HC, Karray F, Lautru S, Gagnat J, Lebrihi A, Huynh TD, Pernodet JL. Glycosylation steps during spiramycin biosynthesis in Streptomyces ambofaciens: involvement of three glycosyltransferases and their interplay with two auxiliary proteins. Antimicrob Agents Chemother. 2010;54(7):2830-2839. . ;:. PubMed Google Scholar
- Song ZQ, Liao ZJ, Hu YF, Ma Z, Bechthold A, Yu XP. Development and optimization of an intergeneric conjugation system and analysis of promoter activity in Streptomyces rimosus M527. J Zhejiang Univ Sci B. 2019;20(11):891-900. . ;:. PubMed Google Scholar
- Cobb RE, Wang Y, Zhao H. High-efficiency multiplex genome editing of Streptomyces species using an engineered CRISPR/Cas system. ACS Synth Biol. 2015;4(6):723-728. . ;:. PubMed Google Scholar
- Balouiri M, Sadiki M, Ibnsouda SK. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016;6:71-79. . ;:. PubMed Google Scholar
- Shirling EB, Gottlieb D. Methods for characterization of Streptomyces species. Int. J. Syst. Bacteriol. 1966;16:312-340. . ;:. Google Scholar
- Frank JA, Reich CI, Sharma S, Weisbaum JS, Wilson BA, Olsen GJ. Critical evaluation of two primers commonly used for amplification of bacterial 16S rRNA genes. Appl. Environ. Microbiol. 2008;74:2461-2470. . ;:. PubMed Google Scholar
- Yoon SH, Ha SM, Kwon S, Lim J, Kim Y, Seo H, Chun J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA and whole genome assemblies. Int. J. Syst. Evol. Microbiol. 2017;67:1613-1617. . ;:. PubMed Google Scholar
- Kieser T, Bibb MJ, Buttner MJ, Chater KF, Hopwood DA. Practical Streptomyces Genetics, John Innes Foundation, Norwich, UK; 2000. . ;:. Google Scholar
- Blaesing F, Mühlenweg A, Vierling S, Ziegelin G, Pelzer S, Lanka E. Introduction of DNA into Actinomycetes by bacterial conjugation from E. coli--an evaluation of various transfer systems. J. Biotechnol. 2005;120:146-161. . ;:. PubMed Google Scholar
- Saleem H, Usman A, Mahomoodally MF, Ahemad N. Bougainvillea glabra (choisy): A comprehensive review on botany, traditional uses, phytochemistry, pharmacology and toxicity. J Ethnopharmacol. 2021;266:113356. . ;:. PubMed Google Scholar
- Berdy J. Thoughts and facts about antibiotics: Where we are now and where we are heading. The Journal of Antibiotics. 2012; 65:385-395. . ;:. PubMed Google Scholar
- Yu Y, Fu Y, Guo X, Yan R, Wang H, Zhao J, Wang X, Zhang J, Xiang W. Streptomyces durbertensis sp. nov., isolated from saline-alkali soil. Int J Syst Evol Microbiol. 2018;68(11):3635-3640. . ;:. PubMed Google Scholar
- Mazodier P, Petter R, Thompson C. Intergeneric conjugation between Escherichia coli and Streptomyces species. J Bacteriol. 1989;171(6):3583-3585. . ;:. PubMed Google Scholar
- Flett F, Mersinias V, Smith CP. High efficiency intergeneric conjugal transfer of plasmid DNA from Escherichia coli to methyl DNA-restricting streptomycetes. FEMS Microbiol Lett. 1997;155(2):223-229. . ;:. PubMed Google Scholar
- Song ZQ, Liao ZJ, Hu YF, Ma Z, Bechthold A, Yu XP. Development and optimization of an intergeneric conjugation system and analysis of promoter activity in Streptomyces rimosus M527. J Zhejiang Univ Sci B. 2019;20(11):891-900. . ;:. PubMed Google Scholar
- Zhang S, Chen T, Jia J, Guo L, Zhang H, Li C, Qiao R. Establishment of a highly efficient conjugation protocol for Streptomyces kanamyceticus ATCC12853. Microbiologyopen. 2019;8(6):e00747. . ;:. Google Scholar
- Rocha D, Ruiz-Villafán B, Manzo M, Rodríguez-Sanoja R, Sánchez S. Development of an efficient conjugal DNA transfer system between Escherichia coli and a non-sporulating Streptomyces strain. J Microbiol Methods. 2018;144:60-66. . ;:. PubMed Google Scholar
- Wang XK, Jin JL. Crucial factor for increasing the conjugation frequency in Streptomyces netropsis SD-07 and other strains. FEMS Microbiol Lett. 2014;357(1):99-103. . ;:. PubMed Google Scholar





