
 Open Access
Open Access Abstract
Metabolic syndrome is a collection of disorders related to metabolisms such as obesity, lipid disorders, hyper/hypoglycemia, ... Metabolic syndrome can lead to cardiovascular diseases, strokes, and diabetes - the leading death causes in the world. In many cases, metabolic disorders are original by the redundant/ reduction of insulin- the most important hormone in metabolism regulation. Both of them are involved in beta-cells dysfunction. Many mechanisms related to this phenomenon has been approved, notably mitochondrial dysfunction and the Ubiquitin proteasome system impairment. UCH-L1 is a protein belonging to the Ubiquitin proteasome system and highly expressed in beta cells. Previous studies reported that decrease UCH-L1 function can alter metabolism and lead to b cell apoptosis under various nutritional conditions, however, the mechanism has not been clarified. In this study, we proposed a Drosophila melanogaster model that expresses many symptoms of metabolic syndrome, by knocking down dUCH (Drosophila homolog of UCH-L1) specifically in Insulin-producing cells. Our fruit fly model had abnormal metabolism, physiology, loss of insulinproducing cells, and mitochondria over-workload, similar to metabolic syndrome in humans. These results suggested that this model is suitable for further studies on the role of UCH-L1 in b cells, as well as a potential model in metabolism diseases' drug screening.
GIỚI THIỆU
Bệnh tiểu đường nói riêng và bệnh rối loạn chuyển hoá nói chung là bệnh mang nhiều triệu chứng của việc rối loạn chuyển hóa như giảm khả năng hấp thu, chuyển hóa carbohydrate, tăng cholesterol trong máu, béo phì,… 1 Hiện nay, cơ chế bệnh vẫn chưa được hiểu rõ và bệnh vẫn chưa có các liệu pháp điều trị triệt để. Do đó, việc xây dựng các mô hình biểu hiện triệu chứng bệnh, đặc biệt là các giai đoạn sớm của bệnh đóng vai trò quan trọng trong việc tìm hiểu cơ chế hình thành bệnh, cũng như sàng lọc các hợp chất và thuốc trong điều trị bệnh.
Sự hình thành các bệnh biến dưỡng có liên quan tới sự rối loạn trong hàm lượng các hormone điều hòa biến dưỡng, trong đó insulin là hormone quan trọng nhất. Sự thiếu hụt hoặc dư thừa insulin thường bắt nguồn từ sai hỏng chức năng của tế bào β- nhóm tế bào sinh tổng hợp loại hormone này 2 . Hiện nay, có nhiều cơ chế liên quan đến rối loạn chức năng tế bào β đã được ghi nhận. Trong số đó, sự suy giảm chức năng của hệ thống Ubiquitin Proteasome (Ubiquitin proteasome system-UPS) và sai hỏng chức năng ty thể được công nhận rộng rãi 3 . Ubiquitin carboxyl terminal hydrolase-L1 (UCH-L1) là một protein thuộc UPS với chức năng chính là tháo, gắn nhãn ubiquitin và ổn định lượng monoubiquitin trong tế bào. UCH-L1 được biểu hiện nhiều ở tế bào β, đồng thời các nghiên cứu in vitro cho thấy giảm biểu hiện UCH-L1 làm tăng hiện tượng apoptosis ở tế bào β trong điều kiện ngộ độc lipid (lipid toxicity) 4 . Trên mô hình chuột, giảm UCH-L1 làm tế bào β nhạy cảm hơn với ngộ độc IAPP (IAPP toxicity) và làm chết tế bào β trong điều kiện này 5 .
Ruồi giấm là mô hình được sử dụng từ lâu trong nghiên cứu chức năng gene và cơ chế bệnh trên người. Đây là mô hình được hiểu rõ về mặt sinh lý và di truyền, có vòng đời ngắn và khả năng sinh sản tốt, do đó cho phép rút ngắn thời gian thí nghiệm và thực hiện nghiên cứu với cỡ mẫu lớn. Bên cạnh đó, ruồi có nhóm tế bào thần kinh sinh insulin (Insulin producing cells- IPCs) có vai trò tương tự như tế bào β ở người, các protein insulin-like peptide 2, 3, 5 (ILP2, 3, 5) tương đồng với insulin. Các quá trình sinh tổng hợp và truyền tín hiệu insulin cũng có tính bảo tồn cao giữa người và ruồi 6 . Nhiều nghiên cứu cho thấy, sự thiếu hụt ILPs và chết IPCs làm ruồi giấm có những triệu chứng tương tự như bệnh biến dưỡng ở người như: tăng đường huyết, suy giảm khả năng tích trữ glycogen, tăng hàm lượng lipid trong cơ thể 7 . Ngoài ra, ở ruồi có protein dUCH (Drosophila ubiquitin carboxyl hydrolase) tương đồng với UCH-L1 ở người.
Từ những cơ sở trên, chúng tôi tiến hành xây dựng mô hình ruồi giấm chuyển gene mang kiểu hình của hội chứng rối loạn chuyển hóa thông qua việc knockdown protein dUCH chuyên biệt tại IPCs và đánh giá các chỉ tiêu sinh lý, năng lượng của cơ thể, cũng như kiểu hình của tế bào.
VẬT LIỆU- PHƯƠNG PHÁP
Các dòng ruồi sử dụng và môi trường nuôi
Các dòng ruồi bố mẹ được duy trì trên môi trường chứa 0.8% agar, 5% sucrose, 5% nấm men, 3% sữa bột tại 25 O C. Dòng ruồi RNAi mang trình tự UAS-dUCH.IR (mã v26468, Vienna Drosophila Resource Center-VDRC) được sử dụng để knockdown dUCH theo cơ chế của RNA can thiệp (RNA interference). Hiệu quả knockdown gene dUCH của dòng ruồi này đã được chứng minh trong các nghiên cứu trước đó 8 . Dòng ruồi RNAi mang trình tự UAS-GFP-IR (kí hiệu 9331, Bloomington Drosophila Stock Center-BDSC) được sử dụng lai tạo dòng đối chứng. Dòng ruồi mang trình tự UAS-mitoGFP (8442, BDSC) được sử dụng để biểu hiện GFP tại ty thể. Các dòng ruồi này được định hướng biểu hiện chuyện biệt tại tế bào thần kinh sinh insulin bằng ILP2-GAL4 (48030, BDSC).
Dòng ruồi knockdown dUCH chuyên biệt tại IPCs là F1 của phép lai 48030 và v26468, mang kiểu gene ILP2-GAL4>UAS-dUCH-IR (viết tắt ILP2>dUCH-IR). Dòng ruồi đối chứng là F1 của phép lai giữa 48030 và 9331, có kiểu gene ILP2-GAL4>UAS-GFP-IR (viết tắt ILP2>GFP-IR). Để quan sát kiểu hình ty thể trên dòng ruồi knockdown dUCH và dòng ruồi đối chứng, chúng tôi tiến hành lai tạo dòng ruồi mang kiểu gene ILP2-GAL4>UAS-mitoGFP, UAS-dUCH-IR và ILP2-GAL4>UAS-mitoGFP, +.
Nhằm ổn định các điều kiện về dinh dưỡng, ruồi thí nghiệm được nuôi trên môi trường tổng hợp Mix Formula 4-24® Instant Drosophila Medium (mã số #173200), với tỷ lệ 1g môi trường: 4ml nước cất từ giai đoạn phôi đến hết thời gian thí nghiệm. Ngoài ra, để đảm bảo hoạt động của hệ thống GAL4-UAS và giảm thiểu các ảnh hưởng lên sinh lý của ruồi, ruồi F1 được nuôi tại 28 O C 9 . Chỉ ruồi đực từ 6-8 ngày tuổi được sử dụng trong thí nghiệm để tránh các tác động do sinh sản.
Đánh giá lượng trehalose, glycogen, triacylglycerol và lipid trong toàn cơ thể
Đánh giá được thực hiện tương tự như quy trình được công bố trước đây của Van Hadel và công sự 10 , 11 . Nhóm ruồi (5con/mẫu) được đồng hóa trong PBS-0.05% Tween 20. Sau đó, carbohydrate, glycogen, lipid được phân tách ra khỏi hỗn hợp tương ứng với các dung môi khác nhau lần lượt là methanol, Na 2 SO 4 10% và chloroform. Carbohydrate được sử dụng để định lượng trehalose thông qua phản ứng với 1N HCl, 1N NaOH và anthrone. Glycogen được định lượng thông qua phản ứng anthrone. Triacylglycerol được định lượng bằng Triacylglycerol Reagent (T2449, Sigma Aldrich) theo hướng dẫn của nhà sản xuất. Lipid được đánh giá qua phản ứng với vanillin. Thí nghiệm được lặp lại 8 nhóm cho mỗi dòng ruồi.
Kiểm tra khả năng kháng đói
Nhóm gồm 10 ruồi đực được chuyển vào ống chứa 1% agar. Ruồi được thay môi trường 12 giờ một lần, ruồi chết được đếm sau mỗi 6 giờ. Thí nghiệm được lăp lại với 10 nhóm ruồi cho mỗi dòng.
Kiểm tra cân nặng của ruồi
Nhóm 10 ruồi đực được gây mê và cân bằng cân phân tích (Sartorius, Đức), sau đó tính khối lượng trung bình cho từng con. Thí nghiệm được lặp lại với 8 nhóm ruồi cho mỗi dòng.
Nhuộm miễn dịch huỳnh quang
IPCs được nhuộm với kháng thể kháng ILP2 (nhận từ giáo sư M.Brown, Athens, GA), chuyên biệt cho IPCs với tỷ lệ 1:450. Hình thái ty thể được quan sát thông qua tín hiệu GFP được nhuộm với kháng thể kháng GFP (#598, MBL, Nhật Bản), tỷ lệ 1:250. Quy trình nhuộm được thực hiện như công bố trước đây của Hiệp và cộng sự 8 . Hình ảnh được chụp thông qua kính hiển vi huỳnh quang ECLIPSE NI-U.
Xử lý dữ liệu
Số liệu được thu nhận và xử lý bằng phần mềm GraphPad Prism 8. Kết quả kiểm tra khả năng kháng đói được xử lý bằng Log-rank test, các kết quả khác được xử lý bằng t-test. Số liệu được trình bày với giá trình trung bình và độ lệch chuẩn.
KẾT QUẢ
Mô hình ruồi k nockdown dUCH tại tế bào thần kinh sinh insulin mang các bất thường trong biến dưỡng và sinh lý cá thể
Hội chứng rối loạn biến dưỡng được đặc trưng bởi sự thay đổi trong các tích trữ năng lượng của cơ thể. Do đó, bước đầu tiên trong quá trình đánh giá mô hình ruồi knockdown dUCH tại tế bào IPCs, chúng tôi tiến hành đánh giá các các chỉ tiêu sinh lý và biến dưỡng của cơ thể ruồi.
Ở ruồi, sau khi ăn, dưới tác động của các Insulin-like peptide (ILPs), các tế bào mô mỡ, mô cơ tiến hành hấp thụ glucose, amino acid, aicd béo từ hemolymph. Tại đây, glucose, amino acid, acid béo được chuyển hóa thành glycogen- dạng tích trữ chính của carbohydrate, triacylglycerol- dạng tích trữ chính của lipid. Một phần glucose được chuyển hóa thành trehalose- dạng đường chính tuần hoàn trong hemolymph, tương tự như glucose trong máu ở người 12 . Do đó, chúng tôi tiến hành đánh giá quá trình biến dưỡng ở ruồi mô hình thông qua ghi nhận các chỉ số về hàm lượng glycogen, trehalose, triacylglyerol và lipid tổng.
Kết quả thực nghiệm cho thấy ruồi knockdown dUCH có các chỉ tiêu biến dưỡng bất thường. Cụ thể, ruồi knockdown dUCH có hiện tượng tăng glycogen ( Table 1 , ****p<0.05), tuy nhiên lại làm giảm mạnh trehalose ( Table 1 , ***p<0.001). Bên cạnh đó, knockdown dUCH tại IPCs, không làm thay đổi lượng TAG ( Table 1 , ns p>0.05 ) , nhưng lại làm tăng đáng kể lượng lipid của cơ thể ( Table 1 , ****p<0.0001).
Song song với việc kiểm tra các chỉ tiêu biến dưỡng của ruồi mô hình, chúng tôi cũng kiểm tra các chỉ tiêu sinh lý chung của cơ thể ruồi, bao gồm cân nặng và khả năng kháng đói. Kết quả cho thấy, khối lượng ruồi mô hình không thay đổi ( Table 1 , ns, p>0.05) nhưng khả năng kháng đói của ruồi tăng so với ruồi đối chứng (∆t=1.9h giờ, *p<0.05, Table 1 )
Ruồi k nockdown dUCH mang kiểu hình ty thể bất thường tại tế bào sinh insulin, IPCs
Sự thay đổi trong biến dưỡng ở dòng knockdown dUCH tại IPCs cho thấy sự rối loạn quá trình tổng hợp và/ hoặc tiết các ILPs- quá trình mà ty thể đóng vai trò quan trọng. Ty thể sản xuất ATP- cung cấp năng lượng cho quá trình tổng hợp và tiết insulin. Bên cạnh đó, nhiều sản phẩm thứ cấp từ hoạt động của ty thể là phân tử tín hiệu cho việc tiết insulin 13 . Do đó, chúng tôi tiến hành đánh giá ảnh hưởng của knockdown dUCH ty thể, bước đầu là đánh giá lên hình thái.
Chúng tôi nhận thấy mật độ ty thể ở dòng ruồi knockdown dUCH có xu hướng tăng so với dòng đối chứng. Điều này thể hiện qua việc ở dòng đối chứng, ở phần lớn diện tích tế bào, ty thể tồn tại với mật độ thấp và chỉ tập trung thành lớp dày tại một vùng nhỏ trong tế bào ( Figure 1 A). Trong khi đó, ở dòng knocdown dUCH tồn tại nhiều vùng mà tại đó ty thể tạo thành lớp dày quanh tế bào. Ngoài ra, ở dòng ruồi này, tại nhiều vị trí, ty thể tập trung với mật độ lớn tạo thành các điểm sáng có cường độ cao ( Figure 1 B). Kết quả này gợi ý sự hoạt động quá tải của ty thể bên trong tế bào.
Figure 1 . Ảnh hưởng của knockdown dUCH lên hình thái ty thể của IPCs. (A): Hình thái ty thể ở dòng đối chứng ILP2>mitoGFP, (B): Hình thái ty thể ở dòng knockdown dUCH ILP2>dUCH-IR, mitoGFP (thước trên hình biểu diễn độ dài 10µm; mũi tên trắng chỉ những vùng có nhiều ty thể tạo thành lớp dày trong tế bào; mũi tên vàng chỉ những vị trí ty thể tồn tại với mật độ cao, tạo thành những đốm sáng có cường độ tín hiệu lớn; mũi tên tím chỉ những vùng ty thể tồn tại với mật độ thấp).

Ruồi k nockdown dUCH có sự suy giảm số lượng tế bào sinh insulin, IPCs
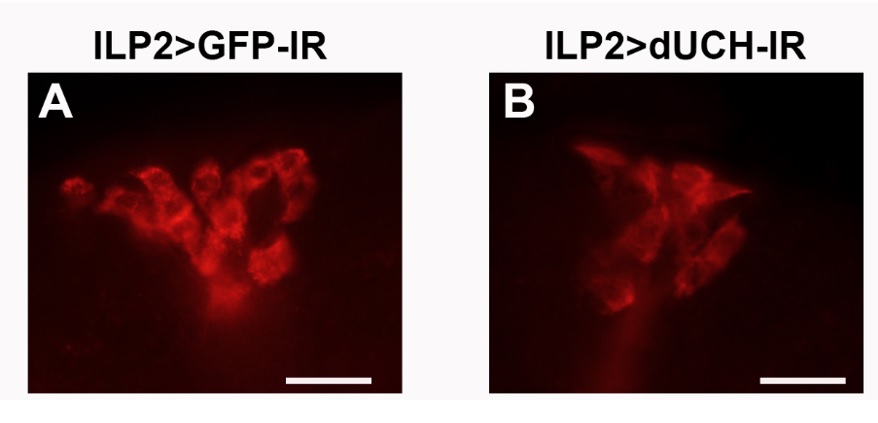
Sự gia tăng ty thể có nhiều khả năng dẫn đến rối loạn chức năng và apoptosis tế bào 14 . Do đó, trong thí nghiệm tiếp theo, chúng tôi tiến hành kiểm tra số lượng IPCs ở các dòng ruồi. Ở ruồi, IPCs tập trung thành cụm 14 tế bào tại não, số lượng IPCs được đánh giá thông qua nhuộm miễn dịch huỳnh quang với kháng thể kháng ILP2. Kết quả cho thấy ruồi knockdown dUCH tế bào có hiện tượng giảm, mất tế bào sinh insulin ( Figure 2 , Table 2 ) .
Figure 2 . Sự suy giảm tế bào sinh insulin ở ruồi knockdown dUCH: Hình nhuộm miễn dịch huỳnh quang đánh dấu IPCs (A) : Dòng đối chứng ILP2>GFP-IR, (B) : Dòng knockdown ILP2>dUCH-IR (thước trên hình biểu diễn độ dài 25µm)
THẢO LUẬN
Các nghiên cứu trước đây cho thấy UCH-L1 đóng vai trò quan trọng trong hoạt động của tế bào β và có liên quan đến bệnh tiểu đường, tuy nhiên cơ chế vẫn chưa được hiểu rõ.
Trong nghiên cứu này, mô hình ruồi knockdown dUCH chuyên biệt tại IPCs của chúng tôi cho thấy những sự rối loạn trong quá trình điều hòa chuyển hóa carbohydrate và lipid, những đặc điểm này có sự tương quan với các hội chứng biến dưỡng ở người ở cả mức độ cơ thể và tế bào ( Table 3 ).
Kết quả về giảm số lượng IPCs trong nghiên cứu của chúng tôi phù hợp với nghiên cứu trước đó của Chu và cộng sự rằng giảm biểu hiện của UCH-L1 làm gia tăng tín hiệu Caspase 3 tại tế bào β trong điều kiện in vitro 4 . Hiện tượng này có thể bắt nguồn từ sự tăng cường hoạt động của ty thể- được thể hiện qua sự tăng tín hiệu bào quan này. Kết quả này đồng thời phù hợp với những thay đổi về mặt biến dưỡng carbohydrate bao gồm: sự tăng hàm lượng glycogen-carbohydrate tích trữ và giảm hàm lượng trehalose- carbohydrate tuần hoàn, đồng thời có thể gợi ý sự tăng hàm lượng ILP2- ILP chính hoạt hóa quá trình hấp thu carbohydrate 13 . Thay đổi trong hoạt động của IPCs cũng là nguyên nhân dẫn tới những rối loạn về biến dưỡng lipid ( Table 1 , Table 3 ). Ngoài ra, sự gia tăng về hàm lượng năng lượng tích trữ dưới dạng glycogen, lipid trong tế bào mặc dù không đủ để cho thấy sự thay đổi về mặt thống kê trên khối lượng, nhưng làm tăng đáng kể khả năng kháng đói của ruồi.
Tóm lại, knockdown dUCH chuyên biệt tại IPCs làm rối loạn hoạt động của ty thể, đồng thời gây suy giảm số lượng tế bào. Đây là nguyên nhân dẫn đến những thay đổi trong quá trình biến dưỡng và sinh lý ruồi. Các kết quả này đồng thời gợi ý tiềm năng của mô hình trong nghiên cứu sâu hơn cơ chế liên quan tới bệnh tiểu đường type 2 và các bệnh biến dưỡng khác.
KẾT LUẬN
Mô hình ruồi knockdown dUCH tại IPCs mang các kiểu hình các hội chứng rối loạn chuyển hóa như tăng tích trữ glycogen, giảm trehalose tuần hoàn, tăng lipid, suy giảm số lượng tế bào sinh insulin và tăng cường hoạt động ty thể. Kết quả nghiên cứu này đóng góp những bằng chứng mới trong cơ chế hoạt động của UCH-L1 trong tế bào β, cũng như mối liên quan của UCH-L1 với bệnh tiểu đường nói riêng và bệnh biến dưỡng nói chung. Kết quả mở ra các nghiên cứu ứng dụng mô hình ruồi knockdown dUCH tại IPCs trong các nghiên cứu cơ chế bệnh tiểu đường, bệnh biến dưỡng và cũng như các nghiên cứu sàng lọc thuốc trị bệnh.
DANH MỤC CÁC TỪ VIẾT TẮT
dUCH: Drosophila Ubiquitin carboxyl hydrolase
IPCs: Insulin-producing cells
ILPs: Insulin-like peptides
ns: not significant
UPS: Ubiquitin proteasome system
UCH-L1 : Ubiquitin carboxyl hydrolase-L1
XUNG ĐỘT LỢI ÍCH
Các tác giả cam đoan rằng họ không có xung đột lợi ích
ĐÓNG GÓP CỦA CÁC TÁC GIẢ
Nghiên cứu này được thiết kế bởi tác giả Đặng Thị Phương Thảo và Đào Mỹ Linh.
Tác giả Đào Mỹ Linh, Huỳnh Mẫn Anh, Võ Thị Hạnh Đan tham gia thu thập số liệu thí nghiệm và xử lý kết quả.
Tác giả Đặng Thị Phương Thảo và Đào Mỹ Linh tham gia viết bản thảo.
LỜI CẢM ƠN
Nghiên cứu này tài trợ bởi Quỹ Nafosted trong khuôn khổ đề tài 108.02-2019.10 và tài trợ một phần bởi Grants-in-Aid trong JSPS Core-to-Core program, B. Asia–Africa Science Platforms.
References
- Hudish LI, Reusch JE, Sussel L. β cell dysfunction during progression of metabolic syndrome to type 2 diabetes. The Journal of clinical investigation. 2019;129(9):. PubMed Google Scholar
- Wilcox G. Insulin and insulin resistance. Clinical biochemist reviews. 2005;26(2):19. Google Scholar
- Muoio DM, Newgard CB. Molecular and metabolic mechanisms of insulin resistance and β-cell failure in type 2 diabetes. Nature reviews Molecular cell biology. 2008;9(3):193-205. PubMed Google Scholar
- Chu K, Li H, Wada K, Johnson J. Ubiquitin C-terminal hydrolase L1 is required for pancreatic beta cell survival and function in lipotoxic conditions. Diabetologia. 2012;55(1):128-140. PubMed Google Scholar
- Costes S, Gurlo T, Rivera JF, Butler PC. UCHL1 deficiency exacerbates human islet amyloid polypeptide toxicity in β-cells: evidence of interplay between the ubiquitin/proteasome system and autophagy. Autophagy. 2014;10(6):1004-1014. PubMed Google Scholar
- Brogiolo W, Stocker H, Ikeya T, Rintelen F, Fernandez R, Hafen E. An evolutionarily conserved function of the Drosophila insulin receptor and insulin-like peptides in growth control. Current biology. 2001;11(4):213-221. Google Scholar
- Haselton A, Sharmin E, Schrader J, Sah M, Poon P, Fridell Y-WC. Partial ablation of adult Drosophilainsulin-producing neurons modulates glucose homeostasis and extends life span without insulin resistance. Cell cycle. 2010;9(15):3135-3143. PubMed Google Scholar
- Tran HH, Dang SN, Nguyen TT, Huynh AM, Dao LM, Kamei K, et al. Drosophila ubiquitin C-terminal hydrolase knockdown model of Parkinson's disease. Scientific reports. 2018;8(1):1-14. PubMed Google Scholar
- Duffy JB. GAL4 system in Drosophila: a fly geneticist's Swiss army knife. Genesis. 2002;34(1‐2):1-15. PubMed Google Scholar
- Handel EV. Rapid determination of glycogen and sugars in mosquitoes. J Am Mosq Control Assoc. 1985;1(3):299-301. Google Scholar
- Handel EV. Rapid determination of total lipids in mosquitoes. J Am Mosq Control Assoc. 1985;1(3):302-304. Google Scholar
- Mattila J, Hietakangas V. Regulation of carbohydrate energy metabolism in Drosophila melanogaster. Genetics. 2017;207(4):1231-1253. Google Scholar
- Semaniuk UV, Gospodaryov DV, Feden'ko KM, Yurkevych IS, Vaiserman AM, Storey KB, et al. Insulin-like peptides regulate feeding preference and metabolism in Drosophila. Frontiers in physiology. 2018;9:1083. PubMed Google Scholar
- Ma ZA, Zhao Z, Turk J. Mitochondrial dysfunction and β-cell failure in type 2 diabetes mellitus. Experimental diabetes research. 2012;:. PubMed Google Scholar
- Tennessen JM, Barry WE, Cox J, Thummel CS. Methods for studying metabolism in Drosophila. Methods. 2014;68(1):105-115. PubMed Google Scholar
- Ellingwood SS, Cheng A. Biochemical and clinical aspects of glycogen storage diseases. Journal of Endocrinology. 2018;238(3):R131-R141. PubMed Google Scholar





