
 Open Access
Open Access Abstract
Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) is the confirmed viral pathogen of COVID-19, a pandemic originated from Wuhan, China at the end of 2019. Since then, SARS-CoV-2 has rapidly spread across the globe with over 8 million confirmed cases and more than 430.000 deaths worldwide as of mid-June 2020. Similar to other strains of coronavirus, the envelope of SARS-CoV-2 comprises of three structural proteins: S protein (spike), E protein (envelope) and M glycoprotein (membrane). SARS-CoV-2 capsids are spherical or pleomorphic. Each capsid contains a positive-sense single-stranded RNA (+ssRNA-Class IV-Baltimore) associated with nucleoprotein N. The viral RNA genome is approximately 30 kb in length and contains 14 open reading frames (ORFs). The binding affinity of the viral S protein to the ACE2 (angiotensin-converting enzyme 2) receptor facilitates the attachment of SARS-CoV-2 to human epithelial cells. Upon binding, SARS-CoV-2 spike protein is cleaved and activated by TMPRSS2 (transmembrane protease, serine 2) or by cathepsin L at the cleavage site S2', and also by furin at the cleavage site S1/S2. The furin cleavage motif RR_R is a notable feature, firstly found in SARS-CoV-2 S protein, which may increase virus transmission rate. This feature and many others might result from several evolution events in SARS-CoV-2 genome. These events could occur when coronaviruses, including SARS-CoV-2, spread from one host to another. They can be causative to high virulence and transmission rate of future coronavirus strains, which may require the development of newer vaccine generations. To understand of SARS-CoV-2’s structure, infection mechanism, diagnosis, treatment, and vaccine development strategies, a review of current literature is of highly importance to disease control in Vietnam.
GIỚI THIỆU
SARS-CoV-2 là tác nhân gây ra đại dịch viêm phổi cấp vào cuối năm 2019 (COVID-19) khởi phát từ Vũ Hán, Trung Quốc. Dịch bệnh ban đầu được nghi ngờ là bắt nguồn từ chợ hải sản Hoa Nam, nơi buôn bán và giết mổ động vật hoang dã lớn của thành phố Vũ Hán, với 27/41 bệnh nhân đầu tiên là công nhân hoặc người có lịch sử dịch tễ liên quan trực tiếp đến khu chợ này. Tuy nhiên, do bệnh nhân đầu tiên không có yếu tố dịch tễ trên nên vẫn chưa có kết luận nào về nguồn gốc khởi phát thật sự của SARS-CoV-2 là thuyết phục tuyệt đối 1 , 2 , 3 , 4 , 5 . Trong lịch sử, các động vật hoang dã như dơi (chi Rhinolophus ), cầy vòi hương ( Paguma larvata C. E. H. Smith, 1827) và lạc đà ( Camelus dromedarius Linnaeus, 1758) là những vật chủ trung gian chứa hàng nghìn loại virus và cũng là thủ phạm gây ra các dịch bệnh do SARS-CoV vào năm 2003 và MERS-CoV vào năm 2012 6 , 7 , 8 , 9 . Tại Đồng Tháp, Việt Nam, chuột và dơi được cho là ổ chứa của hàng trăm loài virus 10 . Phân tích trong một nghiên cứu được công bố vào năm 2018 cho thấy có ít nhất 45 chủng α- và β-coronavirus trong mẫu dơi và chuột 11 . Điều này không chỉ phản ánh tính đa dạng của coronavirus mà còn cho thấy sự nguy hiểm của thói quen tiêu thụ động vật hoang dã ở một số nước trong đó có Việt Nam. Việc xác định nguồn gốc của SARS-CoV-2 vẫn còn nhiều tranh cãi, thậm chí dẫn đến kiện tụng giữa các chính phủ 12 . Trong khi đó, COVID-19 lan khắp thế giới, gây đại dịch với tốc độ lây nhiễm chóng mặt chỉ trong 6 tháng đầu năm 2020 (Đại học Johns Hopkins) 13 , 14 .
Theo ghi nhận vào thời điểm hiện tại (16/06/2020), cả thế giới có hơn 8 triệu ca mắc SARS-CoV-2 ở hơn 200 quốc gia và vùng lãnh thổ, tổng số ca tử vong ghi nhận được là hơn 430.000 ca. Sau khi dịch bùng nổ ở Trung Quốc, các nước có số ca nhiễm SARS-CoV-2 cao là Mỹ, Brazil, Nga, Ấn độ, Tây Ban Nha, Anh, Ý. Trong đó, Mỹ có hơn 2,1 triệu ca nhiễm trong số 8 triệu ca nhiễm của toàn thế giới 13 , 14 . Bộ Y tế Việt Nam ghi nhận 334 ca nhiễm và chưa có bệnh nhân nào tử vong từ đầu dịch đến thời điểm này (16/06/2020) 15 . Mặc dù nguồn tư liệu về bệnh trong giai đoạn đầu là chưa nhiều và chủ yếu được xuất bản bằng tiếng Trung Quốc, tuy nhiên, dữ liệu về COVID-19 tăng nhanh theo thời gian khi dịch bệnh bùng phát sang các nước phương Tây. Việc tổng hợp nguồn tư liệu này là cơ sở để tăng cường hiệu quả kiểm soát dịch bệnh trong nước khi diễn biến tình hình dịch bệnh được dự báo có thể kéo dài đến 2024 16 .
Theo các phân tích sinh học phân tử mới nhất, SARS-CoV-2 đặc biệt thích nghi với việc tấn công gây bệnh trên đường hô hấp ở người qua đường không khí và tiếp xúc với các giọt dịch siêu nhỏ < 10 µm (microdroplet) 17 . Virus mang những biến đổi di truyền độc đáo, trong đó protein S (hay còn gọi là protein gai, protein spike) biến đổi theo hướng sử dụng protease trên tế bào chủ để gắn chặt hơn vào thụ thể, xâm nhiễm và nhân bản hiệu quả hơn. Các biến đổi sinh học của protein S có thể liên quan chặt chẽ đến tốc độ lây nhiễm cao và độc lực tương đối lớn của SARS-CoV-2 trên người. Dữ liệu phân tích di truyền cho thấy SARS-CoV-2 và các virus họ hàng có tốc độ biến đổi di truyền lớn trên protein S 18 , 19 , 20 , 21 , 22 , 23 , 24 , 25 ; điều này tạo ra thách thức lớn đối với việc phát triển và duy trì hiệu quả của các dòng vaccine khi virus có thể đột biến nhanh tạo thành chủng virus có độc lực lớn hơn. Ngoài ra, một số kết quả nghiên cứu cho thấy SARS-CoV-2 có thể tồn tại trong các hạt microdroplet ít nhất ba giờ 26 , 27 , như vậy, liệu điều kiện không khí nhiều bụi có cho phép SARS-CoV-2 phát tán hiệu quả hơn trong điều kiện thực tế ở Việt Nam? Liệu sự khác biệt về đặc điểm di truyền cũng như cấu trúc tháp tuổi của quần thể người châu Á sẽ cho phép Việt Nam có tỉ lệ tử vong thấp như Trung Quốc hay cao như các nước Pháp, Anh, Ý, Tây Ban Nha? Số lượng câu hỏi liên quan đến SARS-CoV-2 là vô tận và sẽ được thu hẹp dần khi tốc độ công bố tăng nhanh. Việc tổng hợp nguồn tư liệu hiện có, hiểu rõ bản chất sinh học, cơ chế xâm nhiễm, phương pháp xét nghiệm, xu hướng điều trị và các chiến lược phát triển vaccine SARS-CoV-2 là tiền đề cấp thiết cho công tác phòng chống dịch hiện nay và cũng là mục tiêu của bài viết này.
MỘT SỐ ĐẶC ĐIỂM DỊCH TỄ HỌC
Các triệu chứng lâm sàng
Người nhiễm SARS-CoV-2 phát bệnh thường có nhiều triệu chứng khác nhau từ nhẹ đến nặng. Sốt, ho và khó thở là những triệu chứng phổ biến nhất chiếm lần lượt 83%, 82% và 31% bệnh nhân. Ở bệnh nhân tiến triển thành viêm phổi thường có hiện tượng xuất hiện các đốm mờ ở phổi khi kiểm tra bằng X quang. Ở bệnh nhân tiến triển thành hội chứng viêm đường hô hấp mạn tính, có sự gia tăng nhanh chóng hiện tượng suy chức năng nhiều cơ quan dẫn đến tử vong. Ngoài ra, khoảng 2-10% bệnh nhân được ghi nhận có các triệu chứng liên quan về đường tiêu hóa như nôn mửa, tiêu chảy và đau bụng 28 , 29 . Tuy nhiên, một số trường hợp được xác định nhiễm virus nhưng không xuất hiện triệu chứng lâm sàng hoặc tiền triệu chứng (pre-symptomic), do đó, những trường hợp này dễ bị bỏ qua dẫn đến số liệu về tỉ lệ người nhiễm virus không triệu chứng vẫn chưa được xác định 30 . Đây là một vấn đề đáng lưu ý trong công tác sàng lọc thông qua triệu chứng để giảm thiểu nguy cơ lây nhiễm trong cộng đồng.
Cách thức virus lây truyền
Hiện nay, cơ chế lây truyền chính xác của SARS-CoV-2 vẫn chưa hoàn toàn được hiểu rõ. Trong đó, việc lây truyền trực tiếp từ người sang người được cho rằng thông qua con đường chủ yếu là tiếp xúc các giọt bắn (droplet) từ người nhiễm virus khi ho, hắt hơi hay nói chuyện 27 . Các giọt bắn có kích thước lớn hơn 10 µm (có khi lớn hơn 1 mm) từ người nhiễm có thể phát tán xung quanh trong khoảng cách 2 m và chỉ có thể gây nhiễm cho người khỏe mạnh khi các giọt này tiếp xúc với họ trước khi rơi xuống đất. Trong khi đó, các giọt bắn có kích thước nhỏ hơn có khả năng lơ lửng và di chuyển xa hơn theo luồng không khí ( Figure 1 ). Nhiệt độ, độ ẩm môi trường là những tác nhân ảnh hưởng đến khả năng lơ lửng của các giọt này 17 , 28 , 31 . Những người khỏe mạnh có thể bị nhiễm bệnh khi tiếp xúc với các giọt bắn trực tiếp từ người nhiễm, hay thông qua tiếp xúc gián tiếp với các đồ vật mang các giọt này. Do đó việc mang khẩu trang và rửa tay thường xuyên là một biện pháp hạn chế tiếp xúc với các giọt bắn thật sự hiệu quả. Ngoài ra, một con đường khác được cho rằng có thể dẫn đến lây nhiễm là đường phân-miệng, từng được ghi nhận ở các trường hợp đối với SARS-CoV và MERS-CoV 31 .

Chỉ số lây nhiễm
Chỉ số lây nhiễm (R0) là chỉ số dùng để chỉ số người có thể bị lây nhiễm từ một người nhiễm virus. Nếu R0 bé hơn 1 thì dịch bệnh có thể được đẩy lùi. Trong khi đó, nếu R0 lớn hơn 1 thì bệnh có thể lây truyền giữa người với người 33 . Dựa trên số liệu của các ca nhiễm ở Trung Quốc thì R0 của SARS-CoV-2 được ước tính khoảng từ 2,24 đến 3,58 34 , ở Ý thì R0 khoảng 2,43 đến 3,10 35 . Trong khi đó, R0 của SARS-CoV là khoảng từ 1 đến 2,75 34 . Giá trị R0 của SARS-CoV-2 cao hơn được cho rằng do thời gian ủ bệnh với các triệu chứng không rõ ràng kéo dài hơn 31 . Hiện nay, thời gian ủ bệnh đối với SARS-CoV-2 là khoảng từ 2 đến 14 ngày, với thời gian ủ bệnh trung bình được ước tính là 5,1 ngày 31 , 36 . Tuy nhiên, con số này có sự thay đổi dựa trên cỡ mẫu quan sát được.
Các yếu tố làm gia tăng nguy cơ tử vong
Một trong những yếu tố làm gia tăng nguy cơ tử vong ở người nhiễm SARS-CoV-2 được ghi nhận rõ nhất hiện nay là tuổi tác. Dựa trên các báo cáo ghi nhận từ tổng số 44.672 ca nhiễm đầu tiên ở Trung Quốc thì tỉ lệ tử vong là khác nhau theo nhóm tuổi ( Table 1 ) 31 , 37 . Người nhiễm có tuổi càng cao thì tỉ lệ tử vong càng cao. Số liệu ghi nhận ở Ý, Tây Ban Nha và Hàn Quốc cũng cho kết quả tương tự, đặc biệt người trên 80 tuổi có tỉ lệ tử vong cao nhất (Ý: 20,2%, ghi nhận ngày 17/03/2020; Tây Ban Nha: 15,6%, ghi nhận ngày 24/03/2020; Hàn Quốc: 13%, ghi nhận ngày 24/03/2020) 38 , 39 . Tuy nhiên, với tình hình dịch ở thời điểm hiện tại (16/ 6/2020), Mỹ là nước có số ca nhiễm cao nhất thế giới (chiếm khoảng 26,9% số ca nhiễm trên toàn cầu) 40 nhưng chưa thể xác định chính xác số người nhiễm tiềm ẩn trong cộng đồng, vì vậy nếu tính tỉ lệ tử vong theo độ tuổi ở Mỹ hay những nước có tình hình tương tự thì cũng không thể hiện chính xác được tỉ lệ này 41 . Do đó, những số liệu về tỉ lệ tử vong theo độ tuổi được công bố chỉ mang ý nghĩa cho thấy yếu tố tuổi có ảnh hưởng tuyến tính đến tỉ lệ tử vong của bệnh nhân COVID-19. Ngoài ra, các yếu tố giới tính và tình trạng bệnh lí nền cũng được ghi nhận có liên quan đến tỉ lệ tử vong của người nhiễm 42 .
| Tuổi | Số ca nhiễm | Tỉ lệ tử vong (CFR) % |
| 0-9 tuổi | 416 | - |
| 10-19 tuổi | 549 | 0,2 |
| 20-29 tuổi | 3.619 | 0,2 |
| 30-39 tuổi | 7.600 | 0,2 |
| 40-49 tuổi | 8.571 | 0,4 |
| 50-59 tuổi | 10.008 | 1,3 |
| 60-69 tuổi | 8.583 | 3,6 |
| 70-79 tuổi | 3.918 | 8,0 |
| Trên 80 tuổi | 1.408 | 14,8 |
ĐẶC ĐIỂM DI TRUYỀN CỦA SARS-CoV-2
SARS-CoV-2 thuộc họ Coronavidae, chi Betacoronavirus , là virus có màng bao (enveloped virus), thông tin di truyền ở dạng RNA mạch đơn, mang mã (mạch dương (+) ssRNA). Vỏ capsid của SARS-CoV-2 không có cấu trúc lập thể cố định (pleomorphism) 43 , chứa phân tử (+) ssRNA khoảng 30 kb (với GC khoảng 38%) có hai vùng trình tự không mã hóa UTR (untranslated region) nằm ở hai đầu, là đặc trưng bộ gene của một -coronavirus ( -CoV), với UTR đầu 5’ dài 265 nucleotide và UTR đầu 3’ dài 229 nucleotide 44 . Trên cơ sở dữ liệu Genbank (NCBI, cập nhật ngày 03/06/2020) có khoảng hơn 90 trình tự bộ gene hoàn chỉnh và hơn 2.700 trình tự bộ gene được lưu trữ trên cơ sở dữ liệu GISSAID (Global Initiative on Sharing All Influenza Data). Các trình tự này thu được thông qua các kỹ thuật giải trình tự Sanger, Illumina, Nanopore và BGI 45 , 46 .
Cấu trúc và đặc điểm trình tự bộ gene của SARS-CoV-2
Bộ gene SARS-CoV-2 chứa các vùng mã hóa chính bao gồm: vùng mã hóa các protein hỗ trợ cho quá trình sao chép của virus (ORF1/ab); vùng mã hóa các protein cấu trúc của virus, gồm protein gai S (spike), protein màng bao E (envelope), protein màng M (membrane) và protein tạo vỏ capsid N (nucleocapsid) ( Figure 2 ) 47 . Trong đó, vùng ORF1/ab được dự đoán mã hóa cho khoảng 16 loại protein khác nhau tham gia vào quá trình sao chép của virus. Trình tự bộ gene của SARS-CoV-2 có sự tương đồng với trình tự bộ gene của các chủng coronavirus (CoV) khác, điển hình là SARS-CoV 48 . Tương tự với các -CoV khác thuộc dòng B, C và D, bộ gene của SARS-CoV-2 không mang vùng trình tự mã hóa cho hemagglutinin-esterase đặc trưng cho các -CoV dòng A ( Figure 2 ) 48 , 49 .
Figure 2 . Các protein cấu trúc đóng vai trò quan trọng trong việc hình thành hạt virus (hình bên trái) 47 . Cấu trúc các vùng mã hóa trên trình tự bộ gene của chủng virus 2019-ncoV (2019 – novel coronavirus, tên gọi mới là SARS-CoV-2) thu nhận từ bệnh nhân Trung Quốc (MN975262) so với các trình tự bộ gene của một số chủng coronavirus như coronavirus ở người (HcoV, human coronavirus) HKU1 (A), SARS-CoV (B), MERS-CoV, CoV HKU9 ở dơi (C) và CoV HKU4 ở dơi (D) (hình bên phải). Vùng 1ab mã hóa các protein tham gia quá trình sao chép bộ gene virus; vùng HE mã hóa hemagglutinin-esterase; vùng S mã hóa protein gai S; vùng E mã hóa protein màng bao E; vùng M mã hóa protein màng M; vùng N mã hóa protein N tạo nucleocapsid; các vùng còn lại mã hóa các protein phụ trợ (accessory protein) 48 .

Trình tự bộ gene của SARS-CoV-2 có sự tương đồng cao với các chủng virus giống SARS. Khi so sánh sự tương đồng về trình tự của bộ gene SARS-CoV-2 cũng như trình tự các vùng mã hóa đều cho thấy SARS-CoV-2 có mối quan hệ họ hàng gần gũi với các chủng virus thuộc chi phụ Sarbecovirus (chi -CoV ) 44 . Đặc biệt, bộ gene của SARS-CoV-2 có trình tự nucleotide tương đồng khoảng 96,2% với bộ gene của chủng virus BatCoV RaTG13 phân lập từ dơi ( Rhinolophus affinis Horsfield, 1823 ) ở tỉnh Vân Nam, Trung Quốc. Trong đó, sự tương đồng về trình tự acid amin của protein S giữa hai chủng này là khoảng 98% 3 , 20 . Ngoài ra, một phân tích của Zhang và cộng sự (2020) còn cho thấy bộ gene của SARS-CoV-2 tương đồng khoảng 91,02% với trình tự nucleotide bộ gene của virus giống SARS từ tê tê ( Manis javanica Desmarest, 1822) 50 . Mặc dù tỉ lệ tương đồng này thấp hơn so với trường hợp chủng virus BatCoV RaTG13 từ dơi, nhưng vùng domain bám thụ thể (RBD) trên protein S ở virus từ tê tê có mức độ tương đồng về trình tự acid amin lên đến 97%, cao hơn 85% ở trường hợp virus từ dơi 51 . Nhìn chung, sự tương đồng cao về trình tự nucleotide và trình tự acid amin với các trình tự ở virus giống SARS được tìm thấy trên động vật cho thấy có khả năng chủng SARS-CoV-2 này có nguồn gốc từ động vật và các quá trình đột biến tái tổ hợp ở bộ gene đã giúp SARS-CoV-2 có khả năng xâm nhiễm lên người 44 .
Cấu trúc protein gai S của SARS-CoV-2
Protein S của SARS-CoV-2 có 1.273 acid amin, gồm hai tiểu phần riêng biệt là S1 và S2, đóng vai trò chủ yếu trong quá trình xâm nhiễm của virus vào vật chủ ( Figure 3 A) . Trong đó, vị trí phân cắt trên protein S (S1/S2) là vị trí xuất hiện mới so với protein S của SARS-CoV cũng như các chủng giống SARS đã biết trước đây, vốn chỉ có một vị trí nhận biết phân cắt là S2’. Vì vậy, protein S của SARS-CoV-2 mang một đặc điểm mới là có đến hai vị trí (S1/S2, S2') để phân cắt S thành S1 và S2 ( Figure 3 A) 20 .
Figure 3 . Cấu trúc chính của protein S ở SARS-CoV-2. RBD: receptor binding domain; RBM: receptor binding motif; TD: transmembrane domain; S1/S2, S2': hai vị trí phân cắt protein S thành S1 và S2. B. So sánh trình tự acid amin trên motif gắn thụ thể (RBM) của SARS-CoV-2 với các chủng giống SARS khác. ACE2 (+): các chủng virus gắn lên thụ thể ACE2; ACE2 (-): các chủng virus không gắn được lên thụ thể ACE2. SARS Fra1: chủng SARS Frankfurt-1; WIV1, WIV16, RsSHC014, Rp3: các chủng từ dơi ở châu Á; BM48-31: chủng từ dơi ở châu Âu 18
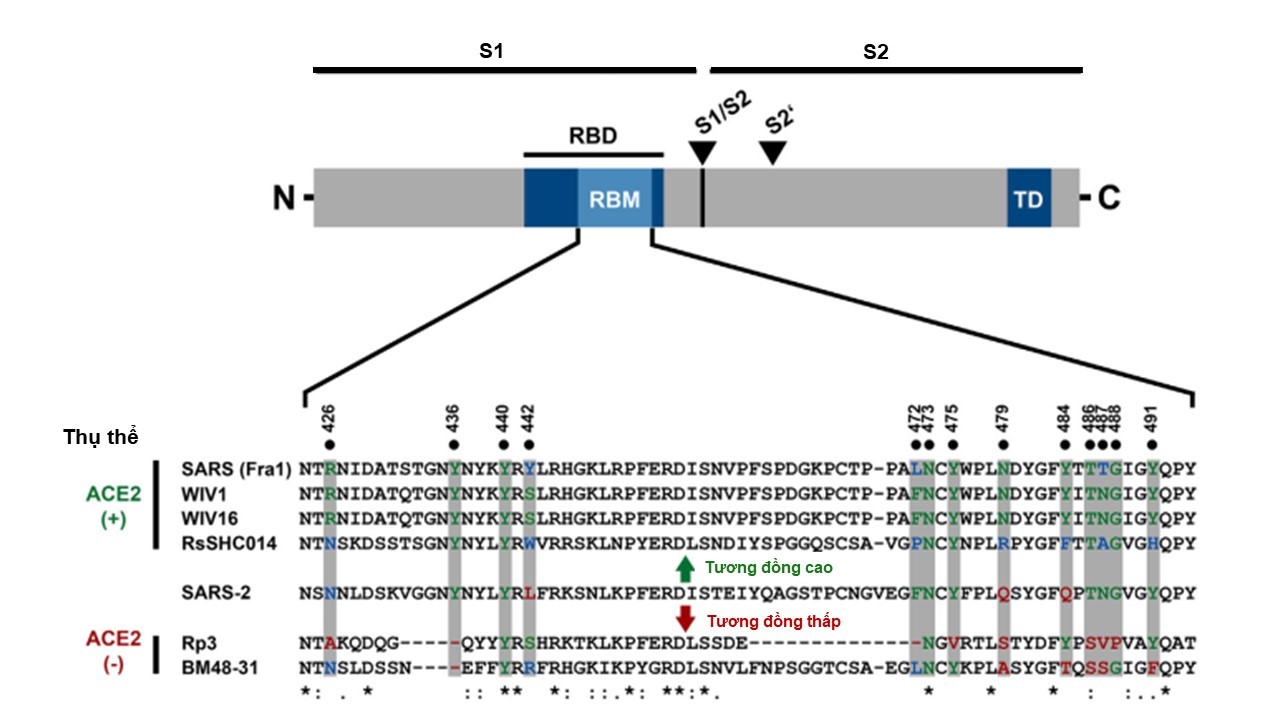
SARS-CoV-2 xâm nhiễm thông qua sự tương tác của protein S với thụ thể ACE2 trên tế bào người. Trong đó, vùng RBD trên tiểu phần S1 của protein S ở SARS-CoV-2 bám vào thụ thể ACE2 của tế bào chủ, giúp virus neo ổn định ở màng tế bào với trạng thái tiền dung hợp và tiểu phần S2 chịu trách nhiệm dung hợp virus vào màng tế bào nhờ bộ máy dung hợp. Ở một số CoV trước đây, việc dung hợp màng sẽ diễn ra khi protein S bị phân cắt tại vị trí S2' nhờ protease cathepsin L hoặc protease TMPRSS2 18 , 19 , 20 , 24 , 52 , 53 . Sự phân cắt này là tín hiệu nhận biết giúp cho protein có hoạt tính dung hợp màng thông qua thay đổi cấu hình 20 . Tuy nhiên, protein S của SARS-CoV-2 mang thêm một vị trí phân cắt mới là S1/S2. Việc sắp gióng cột trình tự acid amin của protein S ở SARS-CoV-2 với trình tự từ các chủng giống SARS cho thấy sự xuất hiện thêm bốn acid amin (P-R-R-A) ở vùng biên giữa S1 và S2 của SARS-CoV-2 đã dẫn đến việc hình thành thêm trình tự nhận biết phân cắt bởi protease furin của người (R-R-X-R, với X là một acid amin bất kỳ) ở SARS-CoV-2 ( Figure 4 ) 19 . Việc xuất hiện motif (P RRAR SV) ở SARS-CoV-2 không được tìm thấy ở các chủng virus giống SARS trước đây, ngay cả ở chủng có độ tương đồng trình tự acid amin ở protein S cao nhất với SARS-CoV-2 là BatCoV RaTG13, ngoại trừ MERS-CoV (với motif P RSVR V) ( Figure 4 b) 50 , 51 , 54 . Tuy nhiên, trong công bố gần đây, một chủng CoV mới (RmYN02) từ dơi ( Rhinolophus malayanus Bonhote, 1903) thu thập ở tỉnh Vân Nam, Trung Quốc có xuất hiện motif P AAR V ở vùng biên giữa tiểu phần S1 và S2. Chủng này được phát hiện có độ tương đồng cao với SARS-CoV-2, độ tương đồng đến 93,3% về trình tự nucleotide bộ gene, chỉ sau BatCoV RaTG13. Trong đó, RmYN02 được xác định là chủng có độ tương đồng về trình tự nucleotide ở vùng 1ab cao nhất với SARS-CoV-2 (97,2%), cao hơn cả trường hợp ở BatCoV RaTG13 (96,5%). Mặc dù độ tương đồng về trình tự acid amin của RBD ở RmYN02 so với SARS-CoV-2 chỉ có 62,4% và có khả năng không sử dụng thụ thể ACE2 để xâm nhập như SARS-CoV-2 nhưng RmYN02 có thể là một bằng chứng đắt giá củng cố cho giả thuyết về nguồn gốc động vật của SARS-CoV-2 55 . Thực tế, một vật chủ có thể tồn tại nhiều chủng CoV, thậm chí các chủng đó thuộc hai chi CoV khác nhau ( α - và β - CoV ) 11 . Do vậy, khả năng xảy ra tái tổ hợp gene giữa các chủng CoV trong vật chủ dẫn đến sự xuất hiện motif trên protein S có chứa trình tự nhận biết phân cắt bởi protease furin là hoàn toàn có thể xảy ra. Tuy nhiên, đột biến này là kết quả của quá trình tái tổ hợp diễn ra trong thời gian rất dài và phải vượt qua được áp lực của chọn lọc tự nhiên để thích nghi với môi trường và vật chủ. Cho đến nay, các nghiên cứu vẫn chưa ghi nhận được chủng CoV ở động vật có trình tự bộ gene giống với SARS-CoV-2. Do đó, nguồn gốc về sự xuất hiện của vị trí nhận biết phân cắt protein S bởi furin ở SARS-CoV-2 vẫn còn nhiều bỏ ngỏ, nhưng rõ ràng đây là một sự xuất hiện không mong đợi.
Figure 4 . Một số đặc điểm nổi bật ở SARS-CoV-2 tại vùng trình tự nucleotide và trình tự acid amin của protein S so với các CoV khác. a. Vùng RBD giúp virus xâm nhiễm vào tế bào chủ thông qua tương tác với thụ thể ACE2. b. Vùng trình tự nhận diện của protease phân cắt giúp cho virus xâm nhiễm vào tế bào chủ 54 .

Walls và cộng sự (2020) đã tiến hành nghiên cứu chứng minh trình tự protein S của SARS-CoV-2 có thể bị phân cắt thành S1 và S2 ngay trong quá trình tổng hợp do tồn tại vị trí nhận biết phân cắt bởi furin. Tác giả cho biểu hiện protein S của SARS-CoV (1.255 acid amin), protein S của SARS-CoV-2 (1.273 acid amin) và protein S ở SARS-CoV-2 bị đột biến (1.269 acid amin) mất vị trí nhận biết phân cắt bởi furin, gọi tắt là mS (mutated S, S đột biến) trong tế bào Vero E6 thông qua hệ thống giả virus MLV (murine leukemia virus). Kết quả kiểm tra biểu hiện bằng Western Blot cho thấy protein S ở SARS-CoV và mS được biểu hiện với kích thước khoảng 180 kDa tương ứng với kích thước của protein S, trong khi đó ở mẫu biểu hiện protein S của SARS-CoV-2 có xuất hiện vạch có kích thước khoảng 90 kDa tương ứng với một trong hai tiểu phần của protein S. Điều này chứng minh rằng protein S của SARS-CoV-2 khi bị đột biến ở vùng nhận biết phân cắt bởi furin (trường hợp mS) thì quá trình biểu hiện sẽ tương tự như protein S của SARS-CoV; trong khi đó, protein S của SARS-CoV-2 bị phân cắt ngay trong quá trình tổng hợp 19 . Bên cạnh đó, một nghiên cứu của Hoffman và cộng sự (2020) cũng chứng minh điều tương tự. Nhóm tác giả cho biểu hiện protein S của SARS-CoV và SARS-CoV-2 tái tổ hợp với hệ thống giả virus VSV (vesicular stomatitis virus) ở dòng tế bào người 293T. Kết quả cho thấy ở mẫu biểu hiện protein S của SARS-CoV có xuất hiện vạch tương ứng với kích thước của protein S (khoảng 180 kDa) nhưng ở mẫu biểu hiện protein S của SARS-CoV-2 lại không xuất hiện vạch tương ứng với kích thước của protein S, mà thay vào đó là vạch tương ứng với kích thước của một trong hai tiểu phần (S1 và S2) của protein S (khoảng 90 kDa) 18 . Như vậy, sự xuất hiện thêm bốn acid amin đã giúp SARS-CoV-2 phân cắt protein S không cần qua trung gian protease cathepsin L hay protease TMPRSS2 mà có thể xảy ra ngay trong quá trình tổng hợp nhờ furin của tế bào chủ 18 , 19 . Hơn nữa, nghiên cứu của Jaimes và cộng sự (2020) cho thấy sự xuất hiện của motif mới trên protein S của SARS-CoV-2 còn cho phép các protease khác không chỉ furin phân cắt protein S thành tiểu phần S1 và S2 như matriptase, cathepsin B và PC1 (proprotein convertase 1) 56 . Điều này có thể có liên quan đến đặc tính lây nhiễm mạnh trong cộng đồng người khi mà cánh cửa tác động đến sự xâm nhiễm của virus vào tế bào chủ thông qua việc phân cắt S1-S2 đã được mở rộng.
Các giả thuyết về nguồn gốc SARS-CoV-2
Hiện nay, vẫn chưa có một cơ sở nào chắc chắn về nguồn gốc chính xác của SARS-CoV-2. Như đã trình bày ở mục 2.1, SARS-CoV-2 có khả năng có nguồn gốc từ động vật, tương tự như các trường hợp SARS-CoV hay MERS-CoV được tìm thấy trước đây có nguồn gốc từ dơi hay lạc đà 6 , 7 , 57 , 58 . Hơn nữa, trình tự bộ gene của SARS-CoV-2 có sự tương đồng cao nhất với trình tự bộ gene của CoV từ dơi BatCoV RaTG13 tính tới thời điểm hiện tại, theo sau đó là RmYN02 cũng có nguồn gốc từ dơi mang motif gần giống với motif mới xuất hiện ở protein S của SARS-CoV-2 51 , 55 . Vì vậy, một số giả thuyết cho rằng SARS-CoV-2 cũng có thể có nguồn gốc từ dơi. Tuy nhiên, giả thuyết dơi là vật chủ trung gian truyền bệnh trực tiếp cho người vẫn chưa được xác thực do các trình tự CoV từ dơi tương đồng với SARS-CoV-2 được thu từ mẫu ở khu vực Vân Nam cách Vũ Hán 1.500 km. Đồng thời, rất ít CoV từ dơi được ghi nhận ở Hồ Bắc và các trình tự CoV này được cho rằng có mối quan hệ khác xa so với SARS-CoV-2 51 . Hơn nữa, bệnh nhân đầu tiên được ghi nhận ở Vũ Hán, Trung Quốc lại không có tiền sử dịch tễ liên quan đến chợ hải sản Hoa Nam 1 . Vì vậy, mặc dù có sự tương đồng về trình tự bộ gene giữa SARS-CoV-2 với CoV từ dơi, nhưng các bằng chứng cho thấy dơi là vật chủ trung gian trực tiếp của SARS-CoV-2 và mối liên hệ với nơi khởi phát dịch đầu tiên là chợ hải sản ở Vũ Hán vẫn chưa rõ ràng.
Một trong những giả thuyết khác về nguồn gốc của SARS-CoV-2 được đưa ra rằng virus này có thể bắt nguồn từ dơi nhưng để có thể lây bệnh trên người thì phải thông qua một vật chủ trung gian khác. Trường hợp này đã được ghi nhận ở SARS-CoV và MERS-CoV với vật chủ trung gian là cầy vòi hương hay lạc đà. Đặc biệt, sự khởi phát đầu tiên của SARS-CoV-2 được cho rằng bắt nguồn từ chợ hải sản ở Vũ Hán, nơi tiêu thụ nhiều động vật hoang dã. Do đó, vật chủ mang SARS-CoV-2 trực tiếp được nghi vấn rằng có thể bắt nguồn từ động vật được buôn bán tại đây. Thông qua việc so sánh trình tự vùng mã hóa giữa SARS-CoV-2 với CoV từ tê tê (thu nhận từ năm 2017 tại Malaysia), Zhang và cộng sự (2020) cho thấy có sự tương đồng trung bình khoảng 93,2% ở cấp độ nucleotide và khoảng 94,1% ở cấp độ acid amin giữa hai chủng này ( Figure 5 ) 50 . Đồng thời, kết quả so sánh trình tự RBD trên protein S cho thấy vị trí này gồm 193 acid amin và tương đồng cao với trình tự RBD ở SARS-CoV-2 từ người (chỉ khác duy nhất 1 aicd amin). Trong khi đó, vùng RBD này giữa SARS-CoV-2 với BatCoV RaTG13 từ dơi khác nhau đến 17 vị trí acid amin và 4 vị trí trong đó quyết định đến khả năng lây nhiễm của SARS-CoV-2. Do đó, CoV từ tê tê được cho rằng có khả năng gây bệnh giống với SARS-CoV-2 ở người 50 . Tuy nhiên, hiện nay vẫn chưa có bất kì cơ sở nào cho thấy tìm được sự hiện diện của SARS-CoV-2 ở tê tê tại chợ hải sản ở Vũ Hán để củng cố cho giả thuyết này. Ngoài ra, không có sự hiện diện của vùng vị trí nhận diện cắt bởi furin trên protein S từ CoV ở dơi hay tê tê ( Figure 4 ), điều này có thể đến từ sự đa dạng vô cùng lớn ở nhóm CoV gây bệnh trên người và động vật nhưng chưa được ghi nhận.
Figure 5 . Kết qua so sánh độ tương đồng trên các gene mã hóa protein giữa CoV từ tê tê (pangolin CoV) với SARS-CoV-2 (người) và BatCoV RaTG13 (dơi) của Zhang và cộng sự (2020) 50 . Độ tương đồng được thể hiện là tương đồng về trình tự DNA/protein.

SỰ TƯƠNG TÁC CỦA SARS-CoV-2 VỚI THỤ THỂ ACE2 – KHỞI ĐẦU CHO QUÁ TRÌNH XÂM NHIỄM
Sự tương tác của SARS-CoV-2 với thụ thể ACE2
Phân tích receptor-binding motif (RBM), là một phần của vùng gắn thụ thể RBD trên protein S của SARS-CoV-2, vùng chịu trách nhiệm tương tác chính với thụ thể ACE2 của tế bào chủ, cho thấy hầu hết các acid amin đều bảo tồn so với RBM của SARS-CoV ( Figure 3 ). Đồng thời, việc so sánh trình tự vùng RBD trên protein S của SARS-CoV-2 với các vùng có chức năng tương tự ở các chủng virus khác cho thấy mặc dù có độ biến động cao nhưng có sự tương đồng từ 75,9% đến 76,9% về trình tự acid amin của RBD giữa SARS-CoV-2 với các chủng virus SARS như SARS-CoV, SARS-like CoV Rs4874, Rs7327, và Rs4231. Các chủng virus này đều có khả năng xâm nhiễm ở người thông qua tương tác với thụ thể ACE2. Trong khi đó, vùng RBD ở các chủng virus SARS gây bệnh ở dơi như Rp3 mang 2 đột biến mất đoạn tại vị trí 433-437 và 460-472, là các vị trí được cho thấy tương tác với thụ thể ACE2, chính vì thế chủng virus này không có khả năng tương tác với ACE2. Ngoài ra, chủng MERS-CoV xuất hiện trước đây tương tác với thụ thể dipeptidyl peptidase-4 (DPP4) để xâm nhiễm mà không phải là thụ thể ACE2 vì vùng RBD không có các acid amin quan trọng cần cho sự tương tác trực tiếp với thụ thể ACE2. Vì vậy, chủng SARS-CoV-2 được cho rằng có khả năng xâm nhiễm ở người thông qua cơ chế tương tác giữa protein S với thụ thể ACE2 nhờ các vị trí acid amin quan trọng, tương tự như các virus SARS gây bệnh ở người 48 , 44 .
Thí nghiệm gây nhiễm SARS-CoV-2 và SARS-CoV lên các dòng tế bào khác nhau từ người của Hoffmann và cộng sự (2020) cho thấy phổ tế bào bị xâm nhiễm bởi SARS-CoV-2 và SARS-CoV là tương đương nhau và đều thông qua sự hiện diện của thụ thể ACE2 trên tế bào chủ ( Figure 6 ) 18 . Ngoài ra, thí nghiệm của Zhou và cộng sự (2020) cho thấy SARS-CoV-2 có thể sử dụng thụ thể ACE2 của người, dơi móng ngựa Trung Quốc, cầy vòi hương và heo được biểu hiện trong tế bào HeLa để xâm nhập vào tế bào nhưng không sử dụng ACE2 của chuột; virus cũng không xâm nhập vào tế bào không có sự biểu hiện của ACE2. Kết quả này minh chứng rằng SARS-CoV-2 thật sự sử dụng thụ thể ACE2 để xâm nhập vào nhiều loại tế bào nhưng không tương thích với thụ thể ACE2 của chuột 3 . Đây là một phát hiện đáng chú ý và có thể gây khó khăn cho các nghiên cứu thử nghiệm liên quan đến SARS-CoV-2 trên mô hình động vật.
Figure 6 . Kết quả gây xâm nhiễm in vitro trên nhiều dòng tế bào người của Hoffmann và công sự (2020) cho thấy phổ tế bào xâm nhiễm của SARS-CoV-2 và SARS-CoV là tương tự nhau 18 .

Nghiên cứu trước đây trên RBD của SARS-CoV cho thấy có năm vị trí acid amin quan trọng trong tương tác với thụ thể ACE2 là Tyr-442, Leu-443, Leu-472, Asn-479, Thr-487 ( Figure 3 B) 59 , 60 . Khi RBD trên protein S của virus bám vào ACE2 trên tế bào người, các vị trí này tạo môi trường kỵ nước và đẩy hai điểm nóng Lys-31 và Lys-353 trên ACE2 quay vào trong hình thành cầu muối (salt bridge) với Glu-35 và Asp-38 trên ACE2, sự hình thành các liên kết cầu muối này cung cấp nguồn năng lượng có lợi cho tương tác giữa RBD của virus với ACE2 của tế bào chủ. Do đó, việc tồn tại của Lys và Ser trên RBD của virus ở cầy vòi hương ( Paguma larvata C. E. H. Smith, 1827), vật chủ trung gian của SARS-CoV trước khi lây qua người, tương ứng với Asn-479 và Thr-487 trên RBD của SARS-CoV, đã làm giảm tính kỵ nước ở vị trí này, từ đó cản trở tương tác giữa RBD và ACE2, khiến nó không tương thích với ACE2 ở người. Nói cách khác, nhờ những thay đổi tại hai ví trí này trên RBD ở SARS-CoV so với virus ở cầy vòi hương, mà SARS-CoV có thể lây nhiễm qua người 59 , 60 , 61 .
Đối với SARS-CoV-2, những vị trí này đã thay đổi theo hướng gia tăng khả năng lây nhiễm, cho thấy chủng virus này dường như đã tiến hóa để đáp ứng có lợi cho tương tác với thụ thể ACE2 ở người. Các vị trí này tương ứng trên RBD ở SARS-CoV-2 đã đột biến thành Leu-455, Phe-456, Phe-486, Gln-493, Asn-501 ( Figure 3 ) 23 , 25 , 61 . Phân tích tương tác của các vị trí đột biến này với thụ thể ACE2 của cầy vòi hương đã ghi nhận Phe-486 hình thành một tương tác bất lợi với chuỗi bên Thr-82 trên ACE2 của cầy vòi hương. Tương tự, Leu-455 và Gln-493 mất tương tác có lợi với ACE2 của cầy vòi hương. Mặc dù vẫn tương ứng được với thụ thể ACE2 của cầy vòi hương nhưng SARS-CoV-2 dường như tiến hóa không theo chiều hướng thích ứng với ACE2 của loài này. Đặc biệt, SARS-CoV-2 không sử dụng thụ thể ACE2 của chuột vì His-353 không tương thích với RBD của virus 61 . Phát hiện này đã ủng hộ bằng chứng về thực nghiệm SARS-CoV-2 không tương tác với thụ thể ACE2 ở chuột của Zhou và cộng sự (2020). Về mặt ý tưởng, mô hình chuột vẫn có thể được sử dụng nếu ACE2 của người được biểu hiện thành công trong chuột. Tuy nhiên, cho đến nay mô hình chuột chuyển gene biểu hiện ACE2 của người vẫn chưa được sử dụng rộng rãi để thử nghiệm vaccine COVID-19 62 . Phân tích vị trí Tyr-442/Leu-455 ở SARS-CoV/SARS-CoV-2 cho thấy Tyr (ở SARS-CoV và virus giống SARS của cầy vòi hương) không mang lại tương tác có lợi với ACE2 và đột biến Tyr thành Leu-455, vốn có tính kỵ nước cao đã giúp tương tác này trở nên có lợi với điểm nóng 31 61 . Ở vị trí Leu-472/Phe-486, cả hai acid amin này đều có lợi trong tương tác với điểm nóng 31 thông qua tương tác kỵ nước với Met-82 của ACE2 ở người. Tuy nhiên, Phe-486 tương tác với Gln-24, Leu-79, Met-82, Tyr-83 của ACE2 ở người trong khi Leu-472 chỉ tương tác với Leu-79 va Met-82 61 , 23 . Tương tự, ở vị trí Asn-479/Gln-493, cả hai acid amin này đều có lợi trong tương tác với điểm nóng 31. Trong đó, Gln-493 hoàn toàn tương thích với điểm nóng 31, tương tác với Lys-31, His-34, Glu-35 của ACE2 ở người trong khi Asn-479 chỉ tương tác với His-34. Ở vị trí Thr-487/Asn-501 thì cả hai acid amin đều tương tác với điểm nóng 353 61 , 23 , 25 . Phân tích này cho thấy các đột biến trong vùng gắn thụ thể RBD của SARS-CoV-2 có liên quan đến tương tác ái lực cao với ACE2 ở người.
Xâm nhập vào tế bào chủ và đáp ứng miễn dịch ở người nhiễm
Trước tiên, SARS-CoV-2 bám được lên màng tế bào người nhờ sự nhận diện và gắn của protein S trên màng bao của virus lên thụ thể ACE2. Tương tự SARS-CoV được phát hiện trước đây, việc xâm nhập vào tế bào niêm mạc của SARS-CoV-2 được cho là cần có sự tham gia của protease xuyên màng trên bề mặt tế bào như TMPRSS2 (Transmembrane protease, serine 2) hoặc cathepsin L: protease xuyên màng xúc tác sự phân cắt và hoạt hóa protein S giúp SARS-CoV-2 đi vào tế bào ( Figure 7 ) 18 , 19 , 31 . Sau khi vào trong tế bào chủ và được giải phóng khỏi màng bao, SARS-CoV-2 tiến hành nhân bản và phiên mã vật liệu di truyền RNA tại tế bào chất. Các protein của virus được tổng hợp ngay tế bào chất sẽ tương tác với nhau và đóng gói RNA bộ gene của virus để tạo thành các hạt virus hoàn chỉnh 31 .
Sau khi xâm nhập vào trong tế bào chủ, SARS-CoV-2 sẽ kích hoạt sự đáp ứng của hệ miễn dịch ở người nhiễm và gây ra chuỗi đáp ứng viêm. Chuỗi đáp ứng viêm này được khởi đầu bằng các tế bào trình diện kháng nguyên từ virus (antigen presenting cell - APC) với hai chức năng chính: (1) trình diện kháng nguyên từ virus cho các tế bào T CD 4 + (Th1) và (2) giải phóng interleukin-12 nhằm kích hoạt Th1. Các tế bào Th1 sẽ hoạt hóa các tế bào T CD 8 + (T killer – Tk) đến tấn công tế bào mang kháng nguyên mục tiêu. Ngoài ra, Th1 còn hoạt hóa tế bào B sản xuất kháng thể kháng lại các kháng nguyên virus 31 , nhờ vậy mà mầm bệnh có thể được tiêu diệt.
Figure 7 . Các bước SARS-CoV-2 xâm nhập vào tế bảo chủ. (A) Bước 1: các protein S trên bề mặt virus tương tác với thụ thể ACE2 trên bề mặt tế bào chủ. (B) Bước 2: phức hợp này được protease xuyên màng (ví dụ TMPRSS2) gắn và phân cắt, đồng thời, protein S được kích hoạt. (C) Bước 3: virus xâm nhập vào tế bào chủ 31 .

KỸ THUẬT XÉT NGHIỆM
Nguyên lý của các kỹ thuật xét nghiệm
Hiện nay, dịch bệnh COVID-19 đã trở thành đại dịch, lan rộng ra toàn thế giới và diễn biến theo chiều hướng khá phức tạp. Do chưa có vaccine phòng ngừa nên cách tốt nhất để ngăn chặn quá trình phát triển của dịch bệnh là sàng lọc các ca nhiễm virus để cách ly với cộng đồng. Do vậy, việc phòng chống dịch hiệu quả không thể chỉ dựa trên chẩn đoán qua những triệu chứng của bệnh nhân mà phải kết hợp với việc xác định bằng các kỹ thuật xét nghiệm dựa trên các phương pháp sinh học phân tử. Dựa trên các đặc điểm sinh học (di truyền và cấu trúc) của SARS-CoV-2, các kỹ thuật xét nghiệm có thể được chia thành hai nhóm, đó là xét nghiệm dựa trên vật liệu di truyền và dựa trên miễn dịch. Mỗi nhóm kỹ thuật này có thể được kết hợp với các kỹ thuật đọc hay hiển thị kết quả khác nhau nhằm rút ngắn tối đa thời gian cho kết quả, đơn giản hóa cách đọc và có thể tự động hoá được.
Các kỹ thuật xét nghiệm dựa trên vật liệu di truyền
Các kỹ thuật xét nghiệm dựa vào PCR
Như đã trình bày trong phần đặc điểm sinh học, vật liệu di truyền của SARS-CoV-2 là RNA nên các kỹ thuật xét nghiệm dựa trên vật liệu di truyền đều dựa trên phương pháp Reverse Transcription-Polymerase Chain Reaction (RT-PCR). Trước tiên, vật liệu di truyền RNA của SARS-CoV-2 được chuyển thành cDNA nhờ phản ứng nghiên mã ngược (Reverse Transcription, RT), sau đó cDNA được dùng làm khuôn để tổng hợp thành DNA mạch đôi và từ đây mạch đôi sẽ làm khuôn cho phản ứng nhân bản chuỗi DNA (Polymerase Chain Reaction/PCR). Đây là các phản ứng cơ bản trong nghiên cứu sinh học phân tử nên vấn đề cần quan tâm ở đây là lựa chọn vùng trình tự mục tiêu để nhân bản. Lựa chọn này ảnh hưởng nhiều tới kết quả của xét nghiệm. Vì SARS-CoV-2 có bộ gene RNA, có tốc độ đột biến cao hơn so với trường hợp bộ gene là DNA 63 , nên cần lựa chọn vùng trình tự mục tiêu ít chịu tác động của tái tổ hợp di truyền nhất. Các bộ kit thương mại hiện nay thường tập trung vào hai vùng chính là ORF1/ab mã hoá cho các protein phi cấu trúc với mức độ bảo tồn cao và vùng đặc trưng cho virus (vùng mã hoá cho protein cấu trúc như protein E hay protein N). Bằng cách kết hợp với mồi/mẫu dò có đánh dấu huỳnh quang, kỹ thuật xét nghiệm RT-PCR có thể giúp nhanh chóng phát hiện, có thể định lượng (RT-qPCR) và có thể tự động hoá. Giới hạn phát hiện thấp nhất hiện nay của xét nghiệm này trong mẫu nghiên cứu là 3,9 bản sao RNA/phản ứng đối với gene mã hóa protein E và 3,6 bản sao đối với gene mã hoá cho protein phi cấu trúc 64 . Một ưu điểm chung của các phương pháp này so với các kỹ thuật xét nghiệm dựa trên miễn dịch là có thể chủ động phát triển bộ kit ở giai đoạn đầu của dịch, khi các sinh phẩm protein liên quan đến mầm bệnh chưa được sản xuất đại trà mà chỉ có thông tin di truyền của mầm bệnh là được công bố rộng rãi.
Các kỹ thuật xét nghiệm đẳng nhiệt
Ngoài kỹ thuật PCR, một số kỹ thuật phát hiện vật chất di truyền khác thuận tiện cho việc đọc kết quả cũng được phát triển như Loop-mediated isothermal amplification (LAMP), Transcription-mediated amplification (TMA) và Recombinase polymerase amplification (RPA). Đây là các phương pháp nhân bản đẳng nhiệt, xét nghiệm dựa trên trình tự acid nucleic, có đặc điểm chung là nhân bản trình tự DNA mục tiêu không cần luân nhiệt (chỉ cần một nhiệt độ cố định). Đặc tính này khiến chúng trở nên phù hợp để triển khai trên phạm vi rộng hơn, ở các điều kiện phức tạp và đa dạng hơn so với kỹ thuật phụ thuộc phản ứng PCR vốn cần nhiều máy móc để thực hiện 64 , 65 , 66 , 67 , 68 .
Về nguyên tắc, kỹ thuật LAMP sử dụng một bộ 4 mồi được thiết kế đặc biệt để nhân bản đặc hiệu DNA thành các cấu trúc “bông cải” giúp đọc kết quả thông qua sự thay đổi màu sắc hay độ đục của sản phẩm phản ứng bằng mắt thường 65 , 66 . Kỹ thuật TMA và RPA đơn giản hơn, chỉ cần sử dụng hai mồi, với thời gian phản ứng ngắn hơn LAMP, chỉ từ 10-30 phút. Độ nhạy và độ đặc hiệu của hai kỹ thuật này tương đương RT-qPCR 67 , 68 , 69 . Tuy nhiên, cần nhấn mạnh là để phát hiện SARS-CoV-2 nói riêng và các tác nhân gây bệnh có bản chất RNA nói chung thì vẫn cần có phản ứng RT diễn ra trước đó. Ngoài ra, do không sử dụng máy luân nhiệt để tách mạch sản phẩm DNA nên cần sử dụng loại polymerase có khả năng tự tách mạch 66 , 70 .
Một phần quan trọng trong việc sử dụng các kỹ thuật nhân bản đẳng nhiệt là việc đọc kết quả. Thông thường người ta có thể cho sản phẩm nhân bản chạy điện di, nhưng việc này đòi hỏi hóa chất và điều kiện của phòng thí nghiệm, làm giảm đi ý nghĩa về sự linh hoạt của phương pháp. Do đó, một số cách thức đọc kết quả ngay tại chỗ (on-site) đã được phát triển, bao gồm chất màu bắt vào DNA dạng mạch đôi như SYBR Green cho tín hiệu của sự tạo thành sản phẩm nhân bản ( Figure 8 ) , hay sử dụng hạt nano vàng để đánh giá sự thay đổi nồng độ dNTP trong phản ứng, qua đó gián tiếp đánh giá sự hiện diện của sản phẩm nhân bản 71 , 72 .
Figure 8 . SYBR green được cho thêm vào sản phẩm phản ứng LAMP cho phép phân biệt mẫu dương tính và âm tính; hình bên phải là mẫu được chiếu bằng đèn tia cực tím 71 .

Kỹ thuật xét nghiệm dựa trên hệ thống CRISPR/Cas
Bên cạnh các phương pháp xét nghiệm truyền thống thì một kỹ thuật mới hiện hành đang rất được quan tâm là xét nghiệm dựa trên CRISPR/Cas. Các protein Cas, mà nổi tiếng nhất là Cas9, về bản chất là các endonuclease/endoribonuclease hoạt động nhờ sự dẫn đường của một đoạn RNA ngắn gọi là guide RNA, có dạng mạch đơn. Protein Cas sử dụng trình tự guide RNA để dò tìm trình tự bổ sung tương ứng trong số các phân tử mà nó bắt gặp. Khi phát hiện ra trình tự mục tiêu, protein Cas thay đổi cấu hình dẫn đến tạo ra đứt gãy trong mạch acid nucleic mục tiêu 73 , 74 . Khi được lập trình để cắt vào sản phẩm nhân bản thì tín hiệu được phát hiện dưới dạng huỳnh quang phát ra nhờ quá trình cắt (tương tự mẫu dò bị phân hủy trong RT-qPCR) và được đọc dưới ánh sáng tia cực tím, hoặc với những mẫu dò thiết kế đặc biệt có thể được phát hiện dưới dạng vạch ở que thử ( Figure 9 ) 75 . Dựa trên nền tảng này, Guo và cộng sự (2020) công bố nghiên cứu xét nghiệm SARS-CoV-2 dựa vào CRISPR/Cas12 và phát hiện nhờ sự phát huỳnh quang qua quan sát dưới ánh sáng xanh dương 76 . Đây được xem là một bước đột phá quan trọng trong các nghiên cứu về kĩ thuật xét nghiệm các virus và có thể sẽ trở thành xu hướng xét nghiệm trong tương lai gần.
Figure 9 . Mẫu chẩn đoán SARS-CoV-2 dựa trên CRISPR/Cas12 ứng dụng kỹ thuật sắc ký miễn dịch dòng ngang để đọc kết quả 77 .

Những ưu điểm lớn của các phương pháp xét nghiệm dựa trên CRISPR/Cas bao gồm: khả năng đơn giản hóa thiết bị mà không phải tốn quá nhiều thời gian (phản ứng nhân bản đẳng nhiệt từ 30 phút – 60 phút, cắt mẫu dò trong khoảng 30 phút, quan sát kết quả huỳnh quang tức thì hoặc chạy trên que thử trong 10 phút) cộng với độ nhạy cao hơn nhiều lần so với phản ứng nhân bản đẳng nhiệt truyền thống nhờ có thêm một lần nhân bản tín hiệu 75 , 76 , 77 . Sự phát triển của phương pháp này hứa hẹn không chỉ là một giải pháp cho vấn đề về phát hiện SARS-CoV-2 mà có thể cho tất cả những tình huống yêu cầu phát hiện DNA hoặc RNA sinh vật một cách đặc hiệu, đơn giản và hiệu quả với chi phí thấp.
Các kỹ thuật xét nghiệm dựa trên miễn dịch
Các kỹ thuật xét nghiệm dựa trên miễn dịch có thể chia thành hai nhóm là xét nghiệm sự hiện diện của virus (kháng nguyên virus) hay xét nghiệm đáp ứng miễn dịch với virus (kháng thể kháng virus). Trong đó, xét nghiệm đáp ứng miễn dịch với virus không thể trực tiếp xác định người dương tính có mang virus hay không, mà chỉ nhằm tìm hiểu xem họ đã từng bị bệnh hay chưa, giúp ích cho công tác tầm soát và nghiên cứu dịch tễ 78 .
Xét nghiệm bằng kỹ thuật Enzyme-Linked Immunosorbent Assay (ELISA)
ELISA là kỹ thuật xét nghiệm thông dụng cho phép xác định sự hiện diện của protein virus trong mẫu bệnh phẩm hoặc kháng thể chống lại virus trong huyết thanh của người được xét nghiệm. Cả hai đều dựa trên nguyên tắc kháng thể đặc hiệu sẽ bắt vào kháng nguyên và nếu một trong hai được cố định trên một bề mặt, thành phần còn lại cũng sẽ bị giữ lại trên bề mặt đó sau bước rửa. Để kiểm tra liệu chất đang quan tâm có bám lên trên bề mặt đó hay không, người ta sử dụng một kháng thể thứ cấp đặc hiệu để phát hiện 78 . Với xét nghiệm sự hiện diện của kháng nguyên SARS-CoV-2 bằng ELISA, các bộ kit thương mại hiện nay dựa trên các kháng thể kháng với kháng nguyên đặc trưng của SARS-CoV-2 như protein S, vùng gắn thụ thể RBD, protein E hay protein N 79 . Bằng cách kết hợp với kháng thể đánh dấu huỳnh quang, nhóm tác giả Diao và cộng sự (2020) có thể phát hiện được protein N của SARS-CoV-2 trong nước tiểu của bệnh nhân sau ba ngày có các triệu chứng điển hình với độ đặc hiệu 73,6% 80 . Tương tự, với xét nghiệm sự hiện diện của kháng thể kháng virus, các bộ kit thương mại hiện nay phát hiện kháng thể đặc hiệu cho kháng nguyên bảo tồn trên protein N 81 của SARS-CoV-2. Bộ kit này có thể phát hiện sự hiện diện của IgA/IgG/IgM sớm nhất là ba ngày sau khi có các triệu chứng điển hình 82 . Kỹ thuật xét nghiệm này cho kết quả hiệu quả nhờ sự đặc hiệu của kháng nguyên và kháng thể, do vậy mà phương pháp ELISA được sử dụng rộng rãi không chỉ trong y học mà còn trong lĩnh vực thực phẩm và nông nghiệp.
Sắc ký miễn dịch dòng ngang - Immunochromatographic Lateral Flow
Để đọc kết quả dễ dàng hơn cũng như có thể tiến hành đại trà ngay tại địa điểm thăm khám (point-of-care), các kit dựa trên kỹ thuật dòng ngang (Lateral Flow Test/LFT) cũng được phát triển. Có thể nói đây là phương pháp xét nghiệm tại chỗ nhanh, rẻ và hiệu quả bậc nhất với đa dạng đối tượng mẫu vật xét nghiệm như nước bọt, mồ hôi, máu và nước tiểu mà một ví dụ điển hình cho phương pháp này là que thử thai. Phương pháp này hoạt động dựa trên nguyên tắc mẫu xét nghiệm sẽ di chuyển dọc theo một dải giấy theo lực mao dẫn, trên đường đi sẽ lần lượt bắt gặp các kháng thể (một và hai) bám đặc hiệu vào protein mục tiêu. Kháng thể một ở dạng tự do nhưng được gắn chất màu, kháng thể hai được cố định vào cellulose trong giấy. Nếu protein mục tiêu hiện diện, nó sẽ tạo nên phức hợp kháng thể hai - protein mục tiêu - kháng thể một tại vị trí mà các kháng thể hai được cố định. Sự tích tụ kháng thể một tại đây sẽ dẫn đến xuất hiện một vạch màu, cho thấy tín hiệu về sự hiện diện của protein mục tiêu 83 . Với 10 µl mẫu huyết thanh sau 10 phút có thể xác định được sự tồn tại của kháng thể IgG/IgM trong mẫu máu với độ đặc hiệu và độ nhạy trên 80% 84 .
So sánh về hiệu quả của các kỹ thuật xét nghiệm
Do mỗi nhóm kỹ thuật được sử dụng để phát hiện các thành phần khác nhau của virus nên hiệu quả, ý nghĩa khoa học - lâm sàng thu được cũng khác nhau. Với nhóm kỹ thuật phát hiện vật chất di truyền, đây là một nhóm kỹ thuật rất nhạy nên về lý thuyết có thể phát hiện tới mức một bản sao, tức một virus/phản ứng và khả năng tự động trả kết quả rất cao khi kết hợp với các phương pháp định lượng sử dụng mồi/mẫu dò có đánh dấu huỳnh quang. Tuy nhiên, cũng chính độ nhạy lại là nhược điểm của kỹ thuật này. Do có thể phát hiện ở nồng độ cực thấp như vậy thì chỉ cần vật chất di truyền của virus tồn tại trong mẫu phân tích chứ không cần nguyên hạt virus thì vẫn có thể cho kết quả dương tính. Điều đó có nghĩa là kể cả trong trường hợp virus không còn khả năng xâm nhiễm nữa thì vẫn cho kết quả dương tính. Do vậy mà một số trường hợp nhiễm COVID-19 ở Việt Nam xảy ra hiện tượng tái dương tính sau khi đã xuất viện. Đây là một lưu ý cần quan tâm khi trả các kết quả lâm sàng.
Với kỹ thuật miễn dịch, hai nhóm kỹ thuật đã trình bày ở trên về bản chất là giống nhau vì đều dựa trên tương tác miễn dịch giữa kháng nguyên và kháng thể nhưng thông tin thu được từ hai kỹ thuật này là hoàn toàn khác nhau. Nếu như kỹ thuật miễn dịch phát hiện kháng nguyên cho biết sự tồn tại của kháng nguyên virus qua đó xác nhận sự hiện diện của virus trong mẫu bệnh phẩm thì kỹ thuật phát hiện kháng thể cho biết cơ thể đã và/hoặc đang tiếp xúc với virus do đáp ứng miễn dịch cần khoảng thời gian từ 3-14 ngày để hình thành kháng thể đặc hiệu. Như vậy, xác nhận sự tồn tại của kháng thể chỉ có giá trị với lần nhiễm bệnh đầu tiên. Với các lần nhiễm thứ hai trở đi thì việc xác nhận này không có ý nghĩa lâm sàng vì không thể phân biệt được kháng thể được tạo ra do lần đáp ứng trước đây hay hiện tại. Do vậy kỹ thuật miễn dịch phát hiện kháng nguyên cho kết quả có ý nghĩa lâm sàng tốt hơn.
| Phương pháp | Reverse Transcription - Polymerase Chain Reaction(RT-PCR) | Reverse Transcription - quantitative Polymerase Chain Reaction(RT-qPCR) | Loop-mediated isothermal amplification (LAMP) | Transcription-mediated amplification (TMA) | Recombinase polymerase amplification (RPA) | Xác định kháng thể |
| Độ nhạy | +++ | +++++ | +++ | ++++ | ++++ | +++ |
| Độ đặc hiệu | ++++ | +++++ | +++ | ++++ | ++++ | +++++ |
| Yêu cầu về thiết bị | +++ | +++++ | + | + | + | + |
| Thời gian phản ứng | ++++ | ++++ | ++++ | +++ | ++ | + |
| Nhiệt độ | 98-55-72 | 98-55-72 | 60 - 65 | 42 | 37 | Ít ảnh hưởng |
| Hiển thị kết quả | Điện di | Tín hiệu huỳnh quang | Điện di, độ đục, màu sắc, tín hiệu huỳnh quang | Điện di, tín hiệu huỳnh quang | Điện di, màu sắc, tín hiệu huỳnh quang | Que thử |
| Thành phần phản ứng | Taq polymerase, reverse transcriptase, 2 mồi, dNTP | Taq polymerase, 2 mồi, reverse transcriptase, 1 mẫu dò, dNTP | 4 – 6 mồi, Bst polymerase, dNTP | 2 mồi, reverse transcriptase, T7 RNA polymerase, dNTP, NTP | 2 mồi, recombin-ase, SSB, DNA polymerase, dNTP | Kháng nguyên SARS-CoV-2, kháng thể anti-human Ig, chất màu |
XU HƯỚNG ĐIỀU TRỊ
Phát triển thuốc
Thuốc ức chế virus mục tiêu
Hiện nay, Tổ chức Y tế Thế giới (WHO), Trung tâm Kiểm soát Bệnh tật Hoa Kỳ (CDC) và Cơ quan Quản lý Thực phẩm và Dược phẩm Hoa Kỳ (FDA) vẫn chưa phê duyệt bất kì loại thuốc đặc trị nào để chữa bệnh COVID-19 rộng rãi 30 , 87 , 88 . Các nhà khoa học đang gấp rút nghiên cứu và phát triển những loại thuốc có khả năng điều trị COVID-19. Phần lớn các loại thuốc điều trị tiềm năng vẫn còn trong giai đoạn thử nghiệm 89 . Việc xem xét sử dụng các loại thuốc kháng virus phổ rộng hiện là một chiến lược kinh tế và hiệu quả trong điều trị COVID-19. Dựa trên thông tin trình tự bộ gene kết hợp với mô hình cấu trúc protein, các nhà khoa học đã đưa ra một danh sách các loại thuốc tiềm năng như: baricitinib, lopinavir-ritonavir, darunavir, favipiravir (favilavir), remdesivir, ribavirin, galidesivir, arbidol, chloroquine, nitazoxanide 90 , oseltamivir (tamiflu), corticosteroid (methylprednisolone), sirolimus, tocilizumab, thiazolidinediones 91 . Trong đó, một số loại thuốc đang được quan tâm như:
Remdesivir
Remdesivir (GS-5734) là một chất tương tự nucleoside adenosine, có khả năng liên kết với polymerase quan trọng của virus là RNA polymerase phụ thuộc RNA (RNA-dependent RNA polymerase, RdRp), ức chế sao chép bộ gene của virus. Remdesivir đã được chứng minh là có hoạt tính kháng các virus phổ rộng, bao gồm Ebola, SARS-CoV và MERS-CoV 92 . Từ các kết quả nghiên cứu trên mô hình động vật và thử nghiệm lâm sàng cho thấy remdesivir là một loại thuốc đầy hứa hẹn trong điều trị COVID-19 93 , 94 . Hiện nay, remdesivir đã bước vào thử nghiệm lâm sàng giai đoạn III ở cả Trung Quốc và Mỹ với nhiều tín hiệu khả quan 90 . Thử nghiệm lâm sàng điều trị COVID-19 (Adaptive COVID-19 Treatment Trial (ACCT)) của National Institutes of Health (NIH) cho thấy thời gian phục hồi trung bình của bệnh nhân sử dụng remdesivir cao hơn 31% (11 ngày) so với bệnh nhân được điều trị bằng hiệu ứng giả dược (placebo) (15 ngày) 95 .
Chloroquine
Chloroquine và hydroxychloroquine là các loại dẫn xuất của 4-amino-quinoline, được dùng trong điều trị sốt rét. Chloroquine và hydroxychloroquine có tác dụng kháng SARS-CoV nhờ vào khả năng tăng pH nội sinh cần thiết cho sao chép bộ gene của virus, cũng như can thiệp vào quá trình glycosyl hóa các thụ thể ACE2 ở tế bào người, giúp ngăn cản sự gắn kết và xâm nhiễm của SARS-CoV-2 89 , 96 . Keyaerts và cộng sự (2004) đã tiến hành thử nghiệm sự ảnh hưởng của chloroquine trên dòng tế bào Vero E6 nhiễm SARS-CoV. Kết quả cho thấy chloroquine có khả năng ức chế 99% khả năng sao chép của SARS-CoV in vitro sau 3 ngày điều trị ở liều 16 μM chloroquine 97 . Hiệu quả ức chế SARS-CoV-2 của hydroxychloroquine được chứng minh là cao hơn so với chloroquine trong một số nghiên cứu in vitro trên tế bào Vero E6. Tại các tỷ lệ nhiễm (MOI) 0,01; 0,02; 0,2 và 0,8 thì nồng độ 50% tác dụng tối đa (EC50) của chloroquine (2,71; 3,81; 7,14 và 7,36 μM) thấp hơn so với hydroxychloroquine (4,51; 4,06; 17,31 và 12,96 μM) 98 . Một số thử nghiệm lâm sàng về khả năng ức chế SARS-CoV-2 của chloroquine và hydroxychloroquine đang được tiến hành. Trong đó, đơn vị nghiên cứu lâm sàng Đại học Oxford, Việt Nam cũng đã đăng ký thử nghiệm lâm sàng giai đoạn 2 cho bệnh nhân COVID-19 tại Việt Nam 99 . Tuy nhiên, hiện tại chưa đủ bằng chứng để kết luận liệu chloroquine và hydroxychloroquine có phải là phương pháp điều trị an toàn và hiệu quả cho COVID-19 hay không 100 .
Lopinavir-Ritonavir
Lopinavir-Ritonavir (LPV-RTV) là một loại thuốc ức chế protease của virus gây suy giảm miễn dịch ở người (HIV) 89 , 101 . Việc điều trị bằng lopinavir-ritonavir (liều uống lopinavir 200 mg/ritonavir 50 mg) làm giảm lượng β-coronavirus trên bệnh nhân COVID-19 xuống mức không phát hiện được 102 . Tuy nhiên, một công bố khác của Cao và cộng sự (2020) về việc điều trị cho 199 bệnh nhân COVID-19 cho thấy quá trình điều trị bằng lopinavir-ritonavir ở những bệnh nhân xuất hiện các triệu chứng lâm sàng nặng không mang lại hiệu quả cao hơn so với điều trị thông thường 101 . Một nghiên cứu khác của Deng và cộng sự (2020) cho thấy thay vì chỉ sử dụng riêng lẽ nhóm LPV-RTV thì việc kết hợp giữa nhóm LPV-RTV và umifenovir (arbidol- một dẫn xuất của indole) đã cho thấy có khả năng tăng tỷ lệ âm tính với SARS-CoV-2 91 .
Favipiravir (favilavir)
Favipiravir là một nucleoside purine có khả năng gây ra sai sót trong tổng hợp RNA của virus. Favipiravir đã được phê duyệt cho thử nghiệm lâm sàng và cho thấy kết quả khả quan trong điều trị COVID-19. Bệnh nhân được điều trị bằng favipiravir cho thấy có sự cải thiện về diễn biến của bệnh (mức độ cải thiện là 91,43% khi quan sát ảnh chụp CT (Computed Tomography) ngực) và khả năng thải loại virus (trung bình sau 4 ngày điều trị) 90 , 103 . Bên cạnh đó, khi so sánh giữa favipiravir với umifenovir trong một thử nghiệm đối chứng ngẫu nhiên cho thấy bệnh nhân COVID-19 được điều trị bằng favipiravir có tỷ lệ hồi phục vượt trội hơn (71,43%) so với điều trị bằng umifenovir (55,86%) và thời gian hạ sốt và giảm ho cũng ngắn hơn đáng kể. Tính đến giữa tháng 4 năm 2020, đã có tám thử nghiệm lâm sàng được thực hiện tại Trung Quốc và Nhật Bản. Các thử nghiệm này nhằm kiểm soát và đánh giá một cách hiệu quả về độ an toàn và tiềm năng chống SARS-CoV-2 của favipiravir 91 .
Mặc dù các loại thuốc đề cập ở trên bước đầu cho thấy có hiệu quả trong điều trị COVID-19, tuy nhiên, cần tiến hành những nghiên cứu với quy mô lớn nhằm tăng tính thuyết phục, cũng như chứng minh độ an toàn cho bệnh nhân.
Thuốc y học cổ truyền của Trung Quốc
Dựa trên thành công khi sử dụng thuốc y học cổ truyền Trung Quốc để điều trị cho bệnh nhân vào hai trận dịch lớn năm 2003 (dịch cúm SARS) và năm 2009 (cúm H1N1), một số loại thảo dược đã được sử dụng rộng rãi trong điều trị COVID-19 tại Trung Quốc 104 , 105 . Trong đó có thể kể đến như: chi Cinchona (canh kina), Glycyrrhiza glabra (cam thảo), Lycoris radiata ( hoa bỉ ngạn, thạch toán), Isatis indigotica (thảo đại thanh), Torreya nucifera ( kaya torreya Nhật Bản hoặc hạt nhục đậu khấu Nhật Bản), Houttuynia cordata ( diếp cá ) , Pelargonium sidoides ( thiên trúc quì ) và Scutellaria baicalensis (hoàng cầm). Các loại thảo dược này có thể được sử dụng riêng lẻ, nhưng phần lớn là chúng được dùng kết hợp với nhau trong công thức chữa bệnh dựa trên các lý thuyết của y học cổ truyền Trung Quốc. Khi sử dụng phương pháp này, bên cạnh yếu tố độ tuổi, tình trạng bệnh của bệnh nhân, một số yếu tố khác cũng cần được cân nhắc như khu vực địa lý trồng cây, thời gian thu hoạch, thời gian bảo quản dược liệu cũng nên được xem xét vì chúng có khả năng ảnh hưởng đến hiệu quả điều trị. Ngoài ra, sự an toàn của bệnh nhân cần được chú ý khi sử dụng công thức từ các loại thảo mộc để chữa trị COVID-19, đặc biệt là khi chúng được sử dụng trong thời gian dài. Hiện tại, chưa có bằng chứng lâm sàng trực tiếp về hiệu quả của các loại dược liệu này trong điều trị COVID-19; các nghiên cứu được báo cáo chủ yếu từ các tài liệu trước đây về phòng chống SARS và cúm H1N1 bằng thuốc y học cổ truyền Trung Quốc. Do đó, các nghiên cứu đáng tin cậy nhằm chứng minh hiệu quả thực sự cần được tiến hành trước khi áp dụng điều trị bệnh COVID-19 104 , 105 , 106 , 107 . Ở Việt Nam, nghiên cứu tinh dầu trong tỏi ( Allium sativum L.) cho thấy 17 hợp chất organosulfur có khả năng tương tác với các acid amin của thụ thể ACE2, hạn chế đường tương tác của SARS-CoV-2 với thụ thể của vật chủ. Đây có thể được xem là một trong số ít các nghiên cứu về các thảo dược có khả năng chống SARS-CoV-2 ở Việt Nam, cho thấy tiềm năng điều trị đáng kể của các thảo dược trong tương lai 108 .
Điều trị triệu chứng và điều trị hỗ trợ
Hiện nay, bệnh COVID-19 vẫn chưa có thuốc đặc trị được phê duyệt chính thức, do đó phương pháp chủ yếu là điều trị triệu chứng lâm sàng và điều trị hỗ trợ 109 , 110 . Tất cả các bệnh nhân phải được cách ly và được phân loại để điều trị tùy theo mức độ nghiêm trọng của bệnh. Đối với trường hợp bệnh nghi nhiễm SARS-CoV-2, bệnh nhân được theo dõi các triệu chứng lâm sàng và được xét nghiệm khẳng định bằng các kỹ thuật sinh học phân tử. Đối với trường hợp đã xác định bệnh COVID-19, bệnh nhân được phân loại và điều trị dựa trên các triệu chứng lâm sàng. Cụ thể, các bệnh nhân xuất hiện các triệu chứng như sốt, ho khan, mệt mỏi, đau cơ và viêm phổi nhẹ được xác định là các ca bệnh nhẹ. Trong trường hợp này, bệnh nhân cần được duy trì chế độ dinh dưỡng, cân bằng điện giải, cân bằng nội môi, kiểm soát sốt và ho, tránh sử dụng thường xuyên kháng sinh và kháng virus như oseltamivir. Sự an toàn và hiệu quả của các loại thuốc này trên bệnh COVID-19 vẫn chưa rõ ràng, việc lạm dụng thuốc có thể gây ra các phản ứng bất lợi nghiêm trọng 110 , 111 , 112 . Các bệnh nhân xuất hiện các triệu chứng như viêm phổi nặng, nhiễm trùng huyết, suy hô hấp cấp tính, sốc nhiễm trùng hay suy đa cơ quan được xác định là các ca bệnh nặng. Trong đó, rối loạn hô hấp là mối đe dọa chính với bệnh nhân và là nguyên nhân chính gây tử vong. Do đó, ở các trường hợp này, việc sử dụng liệu pháp oxy và các phương pháp hỗ trợ hô hấp cần được áp dụng 111 , 113 .
Liệu pháp miễn dịch
Kháng thể đơn dòng (monoclonal antibody, mAb)
Hiện nay, kháng thể đơn dòng áp dụng cho điều trị COVID-19 vẫn chưa được tìm thấy trên thị trường. Tuy nhiên, với sự hiểu biết ngày càng rộng về SARS-CoV-2 sẽ mang lại nhiều hứa hẹn cho việc nghiên cứu kháng thể đơn dòng kháng virus nhằm điều trị COVID-19 trong tương lai. Virus có khả năng bám và xâm nhập vào tế bào chủ nhờ vào sự tương tác giữa protein S và thụ thể ACE2 trên tế bào chủ. Do đó, các protein trên bề mặt virus được coi là mục tiêu chính để nghiên cứu và phát triển kháng thể đơn dòng chống lại chúng ( Figure 10 ) 114 .
Figure 10 . Một cơ chế trung hòa SARS-CoV-2 sử dụng kháng thể đơn dòng gắn với protein virus ở RBD, ngăn virus gắn vào thụ thể ACE2 114
 114" width="300" height="200">
114" width="300" height="200">
Trên thực tế, cả SARS-CoV và SARS-CoV-2 đều có khả năng xâm nhập vào tế bào chủ dựa trên sự tương tác giữa cùng một thụ thể ACE2 của tế bào chủ với protein S của virus, do đó phương pháp trị liệu tiềm năng để ngăn chặn sự xâm nhập của SARS-CoV có thể được ngoại suy để sử dụng cho SARS-CoV-2 115 . Tuy nhiên, trong số ba kháng thể m396, CR3014, CR3022 có khả năng đặc hiệu với SARS-CoV thì chỉ có CR3022 có khả năng liên kết với RBD của SARS-CoV-2. Hơn nữa, CR3022 chỉ liên kết mà không cạnh tranh vị trí gắn vào RBD ở SARS-CoV-2 với thụ thể ACE2. Điều này được giải thích có thể do sự khác nhau về một số acid amin ở đầu C của RBD giữa SARS-CoV và SARS-CoV-2 22 . Do đó, phương pháp này vẫn cần thêm các nghiên cứu thực nghiệm để chứng minh về hiệu quả trung hòa SARS-CoV-2 và khả năng ngăn cản virus gắn vào thụ thể ACE2 ở tế bào chủ của các kháng thể đã được chứng minh đặc hiệu với SARS-CoV trước đó.
Trong số các phương pháp điều trị miễn dịch nhằm ngăn chặn sự xâm nhập của virus, phương pháp sử dụng kháng thể đơn dòng được ưu tiên nhờ vào các ưu điểm như có tính đặc hiệu, độ tinh khiết cao, nguy cơ nhiễm trùng từ mầm bệnh thấp hơn so với điều trị bằng huyết thanh và các chế phẩm immunoglobulin tiêm tĩnh mạch 115 . Tuy nhiên, việc sản xuất kháng thể đơn dòng ở quy mô lớn tốn nhiều công sức, tiền bạc và thời gian. Điều này có thể khắc phục bằng cách xem xét để nhân bản sản xuất kháng thể đơn dòng dựa trên các hệ thống biểu hiện phù hợp như động vật có vú, thực vật và nấm men 114 .
Huyết tương miễn dịch
Huyết tương miễn dịch (convalescent plasma) là huyết tương thu nhận từ bệnh nhân đã khỏi bệnh, đã được sử dụng để điều trị trong hai dịch bệnh là SARS năm 2003 và MERS năm 2012. Các kháng thể trung hòa trong huyết tương miễn dịch có khả năng liên kết và vô hiệu hóa mầm bệnh, bên cạnh đó, các con đường khác qua trung gian kháng thể như kích hoạt bổ sung, gây độc tế bào phụ thuộc kháng thể hay thực bào cũng có thể đóng góp vào hiệu quả điều trị 116 , 117 . Dựa trên điều đó, các nhà khoa học đã tiến hành thử nghiệm sử dụng huyết tương từ các bệnh nhân đã khỏi bệnh COVID-19 để điều trị cho bệnh nhân mắc COVID-19. Trong công bố của Zhang và cộng sự (2020), bốn bệnh nhân nhiễm SARS-CoV-2 có biểu hiện các triệu chứng nặng được điều trị bằng biện pháp chăm sóc hỗ trợ kết hợp với điều trị bằng huyết tương thu nhận từ bệnh nhân đã khỏi COVID-19. Kết quả cho thấy cả bốn bệnh nhân (bao gồm một bệnh nhân mang thai) đã hồi phục và không có phản ứng bất lợi nghiêm trọng liên quan đến truyền huyết tương 118 . Trong một công bố khác của Duan và cộng sự (2020) về sử dụng huyết tương từ người đã khỏi bệnh COVID-19 (liều 200 ml) để điều trị cho 10 trường hợp bệnh COVID-19 nặng cũng cho kết quả khả quan. Cụ thể, sau ba ngày điều trị bằng truyền huyết tương, các triệu chứng lâm sàng được cải thiện nhanh chóng, hàm lượng RNA SARS-CoV-2 giảm dần cho đến mức không thể phát hiện ở cả 10 bệnh nhân (bốn bệnh nhân âm tính sau hai ngày truyền huyết tương, năm bệnh nhân âm tính sau ba ngày truyền huyết tương, một bệnh nhân âm tính sau sáu ngày truyền huyết tương). Bên cạnh đó, các tác dụng phụ nghiêm trọng không xuất hiện ở tất cả các bệnh nhân sau khi điều trị 117 . Từ các nghiên cứu đã công bố cho thấy hiệu quả của việc sử dụng huyết tương miễn dịch trong điều trị bệnh COVID-19. Tuy nhiên, cả hai nghiên cứu đều có một số nhược điểm như các bệnh nhân vẫn được điều trị hỗ trợ bên cạnh truyền huyết tương và thiếu những ghi nhận về sự thay đổi của hàm lượng cytokine trong quá trình điều trị. Hơn nữa, các bệnh nhân được điều trị bằng thuốc kháng virus mặc dù không chắc chắn về hiệu quả điều trị của thuốc sử dụng. Do đó, không thể loại trừ khả năng thuốc kháng virus có thể góp phần vào sự hồi phục của bệnh nhân 117 , 118 .
Việc sử dụng liệu pháp truyền huyết tương điều trị bệnh có một số điểm cần lưu ý. Mầm bệnh tiềm tàng cũng như những rủi to tiềm ẩn có thể tồn tại trong huyết tương và hiện tượng tăng cường miễn dịch phụ thuộc kháng thể có thể gây những ảnh hưởng đến sự an toàn của bệnh nhân. Bên cạnh đó, việc xác định nồng độ kháng thể tối ưu trong huyết thanh cũng như thời điểm truyền huyết tương tối ưu cũng là yếu tố quan trọng cần xem xét để nâng cao hiệu quả điều trị 117 , 119 .
PHÁT TRIỂN VACCINE
Tình hình phát triển vaccine chống virus corona
Một trong những biện pháp phòng ngừa có thể được sử dụng để chủ động kiểm soát dịch bệnh là phát triển vaccine. Vaccine hoạt động dựa trên nguyên tắc đưa một kháng nguyên vào trong cơ thể để gây ra đáp ứng miễn dịch và tạo ra các kháng thể có khả năng trung hòa loại kháng nguyên đó. Kháng nguyên này có thể là một phần của mầm bệnh, hoặc là bản thân mầm bệnh đã bị bất hoạt hay làm yếu đi, hoặc là tiền chất của chúng như DNA hay RNA. Nói cách khác, vaccine dạy cho hệ phòng thủ của cơ thể cách nhận biết và tiêu diệt mầm bệnh thực thụ một cách hiệu quả, ngăn ngừa nguy cơ nhiễm bệnh 120 . Do đó, việc phát triển vaccine được xem là một ưu tiên cấp thiết trong bối cảnh dịch bệnh bùng phát trên toàn thế giới như hiện nay.
Trước đó, thế giới đã ghi nhận sự bùng nổ của một số dịch bệnh có liên quan đến các chủng CoV gây hội chứng suy hô hấp cấp ở người như dịch SARS khởi phát từ Quảng Đông, Trung Quốc năm 2003 121 và MERS khởi phát từ Trung Đông năm 2012 122 . Đến tháng 12 năm 2019, thế giới lại phải đối mặt với dịch COVID-19 khởi phát từ Vũ Hán, tỉnh Hồ Bắc, Trung Quốc 5 . Đây là đại dịch gây nguy hiểm toàn cầu, đã lây lan 215 quốc gia và vùng lãnh thổ với số ca tử vong lên đến hơn 430.000 người tính đến ngày 16/06/2020 5 . Hơn nữa, dịch bệnh này có thể bùng phát trở lại bất cứ lúc nào trong tương lai, vì vậy việc phát triển vaccine thật sự là một biện pháp cấp thiết để phòng ngừa virus gây bệnh COVID-19. Trong cuộc đua tìm vaccine chống lại các chủng CoV và cụ thể cấp bách trong bối cảnh hiện nay là SARS-CoV-2, các nhà khoa học đang nỗ lực nghiên cứu nhiều loại vaccine khác nhau mà phổ biến là vaccine virus bất hoạt, vaccine vector virus, vaccine tiểu phần, vaccine plasmid DNA và vaccine mRNA. Trong đó một số sản phẩm vaccine đã được nghiên cứu thử nghiệm tiền lâm sàng hoặc lâm sàng trên người ( Table 3 ) 123 . Tuy nhiên, cho đến nay vẫn chưa có một loại vaccine nào chống các chủng CoV, bao gồm MERS-CoV, SARS-CoV và SARS-CoV-2, chính thức được WHO và FDA phê duyệt (5,87,121,122).
| Tổ chức nghiên cứu | Loại vaccine | Quốc gia | Tình trạng thử nghiệm |
| Moderna | mRNA (mRNA-1273) | Mỹ | Thử nghiệm lâm sàng giai đoạn 2 |
| Curevac | mRNA | Đức | Thử nghiệm tiền lâm sàng |
| CanSino Biological | Adenovirus (Ad5-nCoV) | Trung Quốc | Thử nghiệm lâm sàng giai đoạn 2 |
| Sinopharm | Virus bất hoạt | Trung Quốc | Thử nghiệm lâm sàng giai đoạn 1-2 |
| Sinovac | Virus bất hoạt | Trung Quốc | Thử nghiệm lâm sàng giai đoạn 1-2 |
| University of Oxford | Vector virus (ChAdOx1) | Anh | Thử nghiệm lâm sàng giai đoạn 2-3 |
| BioNTech/Fosun Pharma/Pfizer | mRNA (3LNP-mRNA) | Liên Minh Châu Âu | Thử nghiệm lâm sàng giai đoạn 1-2 |
| Novavax | Protein tiểu phần (S) | Úc | Thử nghiệm lâm sàng giai đoạn 1-2 |
| Inovio Pharmaceuticals | Plasmid DNA (INO-4800) | Mỹ | Thử nghiệm lâm sàng giai đoạn 1 |
Vaccine virus bất hoạt
Với ưu điểm nhanh, rẻ và đơn giản, vaccine virus bất hoạt là phương án vaccine đầu tiên được cân nhắc khi dịch bệnh vừa mới bùng phát, đáp ứng được yêu cầu cấp bách trong giai đoạn chờ xác định các epitope cụ thể của kháng nguyên có thể gây ra đáp ứng miễn dịch trên vật chủ 124 . Do đó, Sinopharm và Sinovac ở Trung Quốc tập trung vào phát triển vaccine virus bất hoạt để nhanh chóng tìm ra vaccine chống SARS-CoV-2 và những vaccine này hiện đang được thử nghiệm lâm sàng ở giai đoạn 1 – 2 cho thấy tiềm năng đáng kể của chúng 125 , 126 , 127 , 128 , 129 . Bên cạnh đó, vaccine PiCoVacc sử dụng nền tảng virus bất hoạt với β-propiolactone được phát triển bởi Sinovac đã được Gao và cộng sự (2020) công bố có khả năng gây đáp ứng miễn dịch trên chuột, thỏ và khỉ, tạo ra kháng thể vô hiệu hóa SARS-CoV-2 thành công 129 .
Một vài nghiên cứu trước đây đã tạo ra vaccine virus bất hoạt chống SARS-CoV bằng cách bất hoạt virus sống với các tác nhân như formaldehyde, tia UV (ultraviolet light) hoặc β-propiolactone. Kết quả thử nghiệm vaccine virus SARS-CoV bất hoạt trên mô hình chuột và thỏ cho thấy có cảm ứng sinh kháng thể với nồng độ cao 130 , 131 , 132 , 133 . Kháng thể trong huyết thanh thu được từ mô hình thử nghiệm trên chuột và thỏ có ái lực cao với protein S của SARS-CoV, cụ thể là với RBD của tiểu phần S1 130 . Điều này cho thấy RBD có tiềm năng là vùng kháng nguyên chủ yếu của SARS-CoV bất hoạt gây ra đáp miễn dịch trên mô hình động vật 130 . Trong nghiên cứu sau đó, He và cộng sự (2006) tiến hành thu huyết thanh của động vật mô hình sau khi tiêm vaccine virus bất hoạt với β-propiolactone để phân tích các kháng thể đơn dòng (mAb) gắn đặc hiệu với RBD của SARS-CoV bất hoạt. Tuy nhiên, kết quả phân tích cho thấy một số mAb được phát hiện không bắt với các epitope trên RBD ở peptide S tổng hợp của SARS-CoV. RBD ở SARS-CoV bất hoạt có thể đã bị thay đổi cấu hình gấp cuộn do quá trình bất hoạt với β-propiolactone, dẫn dến một số mAb không có hoạt tính với SARS-CoV thực thụ 134 . Đây có thể là một nút thắt cổ chai cho các nghiên cứu phát triển vaccine virus bất hoạt bằng β-propiolactone. Bên cạnh ưu điểm về thời gian và độ khả thi thì mức độ an toàn của vaccine virus bất hoạt vẫn còn nhiều bỏ ngỏ. Ngoài bốn protein cấu trúc quan trọng như protein S, protein E, protein M và protein N thì trình tự bộ gene của SARS-CoV-2 còn mã hóa nhiều protein nhỏ khác nhưng chưa rõ chức năng ( ) 48 . Do vậy, vaccine loại này có thể mang những rủi ro tiềm tàng, gây ra đáp ứng miễn dịch có hại hay phản ứng viêm không mong muốn. Hơn nữa, quá trình bất hoạt virus sống có thể gây nguy cơ nhiễm bệnh cao cho người sản xuất vaccine, hoặc mang khả năng bùng phát dịch cho cộng đồng những người đã tiêm vaccine virus bất hoạt không hoàn toàn 135 , 136 .
Vaccine vector virus
Dựa vào đặc tính dễ lây nhiễm và khả năng nhân lên hiệu quả trong tế bào chủ của virus mà chúng được ứng dụng trong phát triển vaccine như một dạng vector mang kháng nguyên để vận chuyển kháng nguyên đó đi khắp các tế bào, tạo ra sự đáp ứng miễn dịch tăng cường thông qua việc nhân lên liên tục của vaccine vector virus mà không cần thêm liều bổ trợ. Việc phát triển vaccine dựa trên nền tảng vaccine vector virus có thể nhanh chóng đưa đến các thử nghiệm lâm sàng do một số virus được dùng vector thương mại như adenovirus, retrovirus, lentivirus, VSV (vesicular stomatitis virus) đã được chứng minh là an toàn đối với các thử nghiệm trên người 137 , 138 . Đây cũng là một nền tảng vaccine được ứng dụng để phát triển vaccine chống căn bệnh thế kỉ - HIV (human immunodeficiency virus) 139 , 140 . Bên cạnh đó, một số vaccine chống chủng coronavirus trước đây cũng được nghiên cứu dựa trên nền tảng này với bản chất cũng là một vaccine tiểu phần tái tổ hợp do vector virus mang trình tự biểu hiện protein S của các coronavirus 141 , 142 .
Thử nghiệm trên chồn của Weingartl và cộng sự (2004) sử dụng protein S biểu hiện tái tổ hợp trên adenovirus có thể gây đáp ứng miễn dịch. Cụ thể, protein S được biểu hiện trong rMVA (recombinant – modified vaccinia virus Ankara) làm vaccine rMVA-S cho kết quả có gây ra đáp ứng miễn dịch như dự đoán và đi kèm một tác dụng phụ không mong muốn, đó là sự tổn thương trên mô gan của chồn. Mặc dù đây chỉ là nhận định từ quan sát thực nghiệm và nguyên nhân gây viêm gan vẫn chưa được kết luận đầy đủ, nhưng nhóm tác giả cũng đã cảnh báo các nghiên cứu tiếp theo về phát triển vaccine này cần lưu ý về sự liên quan giữa việc gây đáp ứng miễn dịch bằng rMVA-S chống SARS-CoV với sự tổn thương gan và tốt hơn hết là nên có những bằng chứng thực nghiệm chặt chẽ trước khi thử nghiệm trên người 142 .
Các vaccine vector virus chống SARS-CoV-2 được đưa vào thử nghiệm lâm sàng với tốc độ khá nhanh. Điển hình, ChAdOx1 được phát triển bởi đại học Oxford với nền tảng vector là adenovirus từ tinh tinh đang được thử nghiệm lâm sàng giai đoạn 2 – 3 ( Table 3 ) 143 . Tương tự với nền tảng của ChAdOx1, Zhu và cộng sự (2020) công bố vaccine Ad5 (adenovirus serotype 5) được phát triển bởi CanSino Biological và Viện Công nghệ Sinh học Bắc Kinh đã thử nghiệm thành công trên người ở giai đoạn 1 với khả năng gây đáp ứng miễn dịch cực đại ở ngày thứ 28 sau khi tiêm vaccine 144 và đang được thử nghiệm lâm sàng ở giai đoạn 2 ở Trung Quốc 145 . Cả hai vaccine này được dự kiến sẽ hoàn thành các thử nghiệm lâm sàng trong nửa đầu năm 2021, cho thấy vaccine vector virus chống SARS-CoV-2 có mức độ khả thi và độ hiệu quả khá cao, mở ra hi vọng cho các nghiên cứu phòng dịch trong tương lai.
Vaccine tiểu phần
Để phát triển loại vaccine an toàn và hiệu quả thì vaccine tiểu phần được xem là một vaccine mục tiêu tiềm năng. Đặc biệt khi thông tin về các epitope được nghiên cứu rộng rãi, việc xác định epitope có khả năng gây đáp ứng miễn dịch để thu hẹp phạm vi lựa chọn tiểu phần cho việc phát triển vaccine càng trở nên thuận lợi. Trong bốn protein cấu trúc quan trọng của SARS-CoV cũng như SARS-CoV-2 thì protein S, cụ thể là RBD ở tiểu phần S1 được xác định là có chức năng bám vào thụ thể ACE2 của tế bào chủ 18 , 19 , 20 , 146 . Do đó protein S hay tiểu phần S1 của SARS-CoV cũng như SARS-CoV-2 được xem là một ứng cử viên tiềm năng cho việc gây đáp ứng miễn dịch sinh kháng thể trung hòa kháng nguyên 130 , 146 .
Một số nghiên cứu đã thử nghiệm sản xuất protein tiểu phần S1 tái tổ hợp trên mô hình cây cà chua ( Solanum lycopersicum L.) và cây thuốc lá ( Nicotiana benthamiana Domin) để phát triển vaccine chống SARS-CoV 147 , 148 . Đây là một phương pháp sản xuất vaccine không chỉ an toàn mà còn tiết kiệm chi phí vì hệ thống biểu hiện của thực vật khá an toàn, đồng thời có thể sản xuất trên qui mô lớn với mức chi phí lý tưởng hơn so với sản xuất protein bằng lên men tế bào 149 . Pogrebnyak và cộng sự (2005) đã biểu hiện thành công protein S trong E. coli Rosetta-2 (DE3), cây thuốc lá và cây cà chua. Kết quả thử nghiệm khả năng đáp ứng miễn dịch cho thấy có kích thích phản ứng miễn dịch đối với mô hình chuột cho ăn vật liệu quả cà chua. Trong khi đó, đáp ứng miễn dịch không được phát hiện ở mô hình chuột thử nghiệm bằng cách cho dịch chiết rễ cây thuốc lá thông ống vào đường dạ dày nhưng lại được phát hiện khi tiêm liều bổ sung peptide S thương mại 148 . Tuy nhiên, nghiên cứu này vẫn chưa cho thấy liệu các kháng thể từ các thử nghiệm miễn dịch này có khả năng vô hiệu hóa SARS-CoV hay không cũng như hoạt tính thực thụ của vaccine sản xuất bằng mô hình này đối với SARS-CoV vẫn chưa thể hiện rõ. Điều này cho thấy sản phẩm protein tái tổ hợp S1 của SARS-CoV được tạo ra ở thực vật có thể gây đáp ứng miễn dịch không cao bằng so với các phương pháp khác. Tuy nhiên, nếu phương pháp này được cải tiến về mặt sản lượng cũng như cấu trúc của protein tái tổ hợp để tăng khả năng đáp ứng miễn dịch thì đây có thể là một bước đột phá quan trọng trong lĩnh vực phát triển vaccine không cần tiêm chủng như truyền thống, mà thay vào đó là được bổ sung trực tiếp qua đường tiêu hóa. Mặc dù phát triển vaccine tiểu phần vẫn còn nhiều rào cản về tốc độ thời gian thử nghiệm nhưng dữ liệu ghi nhận ngày 03/06/2020 cho thấy vaccine tiểu phần chống SARS-CoV-2 do Novavax phát triển đã bước vào thử nghiệm lâm sàng ở giai đoạn 1-2. Nghiên cứu thử nghiệm dự kiến sẽ hoàn thành vào cuối tháng 7/2021, cho thấy tiềm năng rất lớn của nền tảng vaccine này 150 .
Vaccine DNA và mRNA
Khác với các vaccine có bản chất là protein của mầm bệnh, vaccine DNA mang vật liệu di truyền mã hóa cho mầm bệnh được thiết kế ở dạng plasmid. Plasmid này chịu sự kiểm soát của promoter ở virus như cytomegalovirus (CMV) giúp nó biểu hiện hiệu quả nhờ hệ thống di truyền của vật chủ. Protein được mã hóa bởi DNA này có thể được cơ thể vật chủ nhận biết như mầm bệnh virus để trình diện lên bề mặt tế bào, kích thích phản ứng miễn dịch 151 , 152 , 153 . Đây được xem là nền tảng vaccine hiệu quả về mặt sản xuất vì plasmid DNA dễ dàng được tăng sinh trong tế bào vi khuẩn với mức chi phí rất lý tưởng. Do đó, sau khi dịch SARS (2003) bùng nổ, có khá nhiều vaccine DNA được thiết kế cho kết quả khả quan trên mô hình động vật và thử nghiệm lâm sàng trên người thành công ở giai đoạn 1 152 , 154 , 155 . Vaccine DNA cũng là một ứng cử viên tiềm năng trong cuộc đua tìm vaccine chống SARS-CoV-2 khi INO-4800 (vaccine DNA được phát triển bởi INOVIO Pharmaceuticals) đã bước vào thử nghiệm lâm sàng giai đoạn 1 156 . INO-4800 nhắm mục tiêu vào protein S của SARS-CoV-2 và được chèn thêm trình tự dẫn đường IgE thành một trình tự tổng hợp. Trình tự này được tối ưu hóa codon để có thể biểu hiện trong tế bào người, sau đó dòng hóa vào vector pGX0001 có sẵn promoter CMV và hormon điều hòa tăng trưởng bovine kết thúc tín hiệu phiên mã. Thử nghiệm vaccine này trên mô hình động vật cho kết quả rất khả quan, có khả năng gây đáp ứng miễn dịch và ức chế tương tác giữa thụ thể ACE2 với protein S của SARS-CoV-2 trong huyết thanh của động vật thử nghiệm 151 . INO-4800 được đưa vào thử nghiệm lâm sàng giai đoạn 1 từ đầu tháng 4/2020 và dự kiến sẽ hoàn thành thử nghiệm lâm sàng vào tháng 4/2021 156 . Đây không chỉ là một tín hiệu khả quan cho vaccine chống SARS-CoV-2 mà còn là niềm hi vọng rất lớn cho sự phát triển của nền tảng vaccine DNA.
Các phân tử mRNA kém bền, dễ bị phân hủy bởi RNase nhưng khi được bảo vệ, chúng lại trở thành dạng vaccine đột phá trong việc phòng và trị bệnh. Phân tử mRNA sau khi tổng hợp trong điều kiện in vitro được chuyển vào trong tế bào người. Phân tử này sẽ sử dụng bộ máy dịch mã trong tế bào đích để biểu hiện sản phẩm được mã hóa và các protein này gây ra đáp ứng miễn dịch. Báo cáo đầu tiên về việc sử dụng mRNA như một dạng vaccine là việc biểu hiện thành công nucleoprotein (NP) của virus cúm, kích hoạt tế bào T gây độc chuyên biệt NP 157 . Sau đó, vaccine mRNA đã tiếp tục gặt hái được nhiều thành tựu ở lĩnh vực miễn dịch học trong việc phòng chống/điều trị các tác nhân gây bệnh như virus Zika 158 , virus dại 159 , cytomegalovirus 160 và cả tế bào ung thư 161 với độ an toàn và tính hiệu quả cao.
Vaccine mRNA thể hiện nhiều ưu điểm vượt trội so với các dạng vaccine khác. So với DNA, phân tử mRNA kém bền hơn và sẽ bị phân hủy đáng kể trong tế bào sau vài ngày 162 . Nhờ vậy, vaccine mRNA thực hiện chức năng của một chất kích thích miễn dịch tạm thời và không ảnh hưởng lâu dài đến tế bào. Ngoài ra, mRNA không cần phải đi vào nhân mà hoạt động ở tế bào chất, không gây ảnh hưởng đến cấu trúc bộ gene nên nó không gây hại cho tế bào về mặt di truyền và cũng không tự nhân đôi. So với các vaccine có bản chất peptide/protein, mRNA mang trình tự cần biểu hiện được dịch mã chính xác trong tế bào người nên giúp loại bỏ các trở ngại khi biểu hiện peptide/protein tái tổ hợp ở các sinh vật khác trong quá trình sản xuất. Đối với các mầm bệnh có khả năng đột biến mạnh, tạo ra nhiều biến chủng, trình tự của mRNA vaccine dễ dàng thay đổi các nucleotide mà không gây ảnh hưởng nhiều đến đặc điểm lý hóa. Petsch và cộng sự (2012) chứng minh vaccine mRNA là liệu pháp hiệu quả để đối phó với khả năng đột biến lớn của virus cúm khi đưa ra các bằng chứng về khả năng kháng virus cúm ở động vật 163 .
Quá trình sản xuất vaccine mRNA được thực hiện trong điều kiện in vitro và cần được tối ưu hóa để đạt được hiệu suất biểu hiện cao khi chuyển vào điều kiện in vivo . Phần lớn phân tử mRNA được hình thành dựa trên sự phiên mã của một phân tử DNA có promoter phù hợp với RNA polymerase của thực khuẩn thể, một ORF và một trình tự mã hóa cho đuôi poly(A). Hỗn hợp sản phẩm sau đó sẽ được xử lý với DNase để loại bỏ DNA. Phân tử mRNA được tổng hợp in vitro dưới sự xúc tác của RNA polymerase thực khuẩn thể cần được bổ sung thêm mũ 7-methyl guanosine (7 m G) 164 ở đầu 5’ hoặc 3´O-me 7-meGpppG (tên thương mại là Anti-Reverse Cap Analog, ARCA) để tăng hiệu suất dịch mã và đuôi poly(A) ở đầu 3’ 165 để đảm bảo sự dịch mã xảy ra hiệu quả trong tế bào eukaryote 166 . Việc biến đổi sau phiên mã này được thực hiện sau khi phản ứng polymer hóa kết thúc, trong một số trường hợp, m 7 G (5’)-ppp-(5’) G có thể được bổ sung với lượng dư vào phản ứng polymer hóa để nó có thể là nucleotide đầu tiên bắt đầu quá trình phiên mã. Vì được biểu hiện trong tế bào động vật, codon mở đầu của mRNA cần có trình tự tương ứng với trình tự Kozak để tăng hiệu suất dịch mã 167 . Các vùng không dịch mã (untranslated region, UTR) khi được tối ưu hóa cũng có khả năng tăng cường biểu hiện của mRNA 168 , 169 . Phân tử mRNA sau khi sinh tổng hợp và biến đổi sẽ được tinh sạch bằng phương pháp sắc kí 170 và được chuyển vào tế bào thông qua một số phương pháp như dùng xung điện với mRNA trần hoặc kết hợp với nhiều loại chất mang khác như protamine 163 , liposome-protamine 171 , hạt nano lipid 158 , 160 ( Figure 11 ).
Figure 11 . Minh họa một số phương pháp phổ biến đưa phân tử mRNA vào tế bào. Phân tử mRNA tổng hợp in vitro được kí hiệu bằng một đường liền có mũi tên chỉ trình tự của mRNA, theo chiều 5’-3’; bắt đầu bằng mũ 7-methyl guanosine (7mG) và kết thúc bằng trình tự polyA. a) Dùng xung điện để đưa mRNA trần vào tế bào. b) Dùng chất mang protamine (được kí hiệu ở dạng khối sáu thùy), là các protein giàu Arginine giúp dễ dàng liên kết với mRNA. c) Dùng chất mang protamine và màng liposome: liposome cấu tạo bởi lớp đôi phospholipid bao quanh phức hợp protamine-mRNA. d) Dùng các hạt nano lipid để đưa mRNA vào tế bào: lớp đôi phospholipid và các phân tử cholesterol hình thành lớp màng bao bọc phân tử mRNA.

Box
Moderna (Mỹ) và CureVac (Đức) là hai công ty vaccine hàng đầu thế giới đang sử dụng nền tảng mRNA để sản xuất nhiều loại vaccine trong đó có COVID-19. Moderna công bố tiến hành thử nghiệm vaccine mRNA COVID-19 trên người ngay sau khi dịch bệnh bùng nổ cho thấy công nghệ mới này đang khẳng định được tầm ảnh hưởng lẫn độ hiệu quả. Tương tự như Moderna, CureVac được thành lập vào năm 2000, là một công ty với khoảng 200 bằng sáng chế tập trung cho vaccine mRNA, nhiều loại trong số đó đang thử nghiệm giai đoạn 1. Ngày 3/3/2020, Daniel Menichella, CEO tại Mỹ của CureVac đã có cuộc gặp với tổng thống Mỹ với tư cách thành viên nhóm phản ứng coronavirus của nhà Trắng (White House Coronavirus Task Force). Cuộc gặp đã đề cập đến một thương vụ nhằm chuyển công nghệ từ Đức đến Mỹ, sản xuất độc quyền vaccine COVID-19 cho người Mỹ. Việc này dẫn đến thông báo phản đối của chính phủ Đức và Daniel Menichella bị thay thế vào ngày 12/3. Ngay sau đó, ngày 16/3/2020, Liên minh châu Âu EU kí khoản hỗ trợ 80 triệu euro cho CureVac nhằm tập trung nghiên cứu sản xuất vaccine COVID-19. Câu chuyện về sự tranh giành CureVac, mà bản chất là tranh giành công nghệ sản xuất vaccine mRNA giữa hai cường quốc không chỉ cho thấy sức ảnh hưởng của COVID-19 mà còn thể hiện sức mạnh tuyệt đối của công nghệ sinh học trong điều chế thuốc và phòng chống bệnh dịch.
KẾT LUẬN
SARS-CoV-2 là một (+) ssRNA khoảng 30 kb với bốn protein cấu trúc chính đóng vai trò hình thành nên hình dạng của virus là protein S, E, M, N. Trong đó, protein S chịu trách nhiệm chính trong quá trình xâm nhiễm vào tế bào chủ. Hiện nay dữ liệu bộ gene của SARS-CoV-2 được công bố khá rộng rãi trên NCBI và GISAID. Dựa vào dữ liệu đó, nghiên cứu về cấu trúc và chức năng của các protein S, E, M, N cũng như nghiên cứu về thông tin di truyền của SARS-CoV-2 dần được hoàn thiện. Tuy nhiên, nguồn gốc của SARS-CoV-2 vẫn đang là dấu chấm hỏi lớn chưa có lời đáp, cho thấy mối đe dọa về virus này vẫn đang tiềm ẩn.
Hiện nay, ngoài các kỹ thuật xét nghiệm truyền thống dựa trên PCR thì các kỹ thuật xét nghiệm mới với nhiều ưu điểm vượt trội đang dần chiếm lĩnh thị trường như xét nghiệm đẳng nhiệt và xét nghiệm dựa trên CRISPR/Cas. Các kỹ thuật xét nghiệm này cho phép xác định trong thời gian ngắn và có thể thực hiện linh hoạt mà không bị giới hạn về điều kiện thiết bị cồng kềnh và chi phí đắt đỏ. Đây đều là các kỹ thuật phần nào khẳng định vị thế của công nghệ sinh học phân tử trong hầu hết các lĩnh vực và dự đoán sẽ còn phát triển xa hơn nữa trong tương lai.
Cuộc đua tìm thuốc điều trị và vaccine chống các chủng coronavirus tại thời điểm thực hiện bài viết tổng quan này (khoảng 6 tháng từ khi dịch COVID-19 bùng phát) vẫn chưa cho thấy có dấu hiệu hạ nhiệt và cũng chưa có hồi kết. Tuy nhiên, một số sản phẩm thuốc và vaccine chống SARS-CoV-2 đã được tiến hành thử nghiệm lâm sàng với tốc độ nhanh chóng chưa từng có. Điều này gợi ra hy vọng thế giới sẽ sớm có thuốc điều trị và vaccine phòng ngừa. COVID-19 vẫn đang diễn biến phức tạp ở Mỹ, các nước Nam Mỹ, Nga, Ấn độ và nhiều nước khác; trong khi đó tình hình dịch ở Việt Nam có thể được xem như đã hạ nhiệt nhờ thực hiện giãn cách xã hội hiệu quả. Dự đoán thiệt hại về kinh tế và xã hội do COVID-19 để lại có thể tồn đọng trong khoảng thời gian khá dài. Do đó, mọi nỗ lực phát triển thuốc và vaccine vẫn đang được tập trung không chỉ để giải quyết cuộc khủng hoảng về sức khỏe và y tế trong bối cảnh hiện tại mà hướng đến ngăn chặn sự tái bùng phát dịch trong tương lai.
Các thông tin về đặc điểm của SARS-CoV-2 là tiền đề cho các nghiên cứu sâu hơn về kĩ thuật xét nghiệm, xu hướng điều trị và phát triển vaccine. Mặc dù những dữ liệu này còn nhiều khoảng trống vẫn chưa được khám phá, tuy nhiên các phát hiện quan trọng được đề cập trong bài viết này đã củng cố phần nào những hiểu biết về SARS-CoV-2. Hơn nữa, dữ liệu thực nghiệm về SARS-CoV-2 liên tục được công bố và ngày càng chi tiết, hỗ trợ một cách đắc lực cho công tác phòng chống và điều trị COVID-19.
DANH MỤC CÁC TỪ VIẾT TẮT
ACE2: Angiotensin–converting enzyme 2
APN: Aminopeptidase N
CFR: Case fatality rate
CoV: Coronavirus
COVID-19: Coronavirus disease – 2019
CDC: Centers for Disease Control and Prevention
DPP4: Dipeptidyl peptidase–4
ELISA: Enzyme–Linked Immunosorbent Assay
FDA: Food and Drug Administration
HE: Hemagglutinin-esterase
LAMP: Loop-mediated isothermal amplification
mAb: Monoclonal antibody
MERS-CoV: Middle East respiratory syndrome coronavirus
mS: Mutated S
NCBI: National Center for Biotechnology Information
NP: Nucleoprotein
ORF: Open reading frame
PCR: Polymerase chain reaction
RT – PCR: Reverse transcription – Polymerase chain reaction
RT – qPCR: Reverse transcription – quantitative Polymerase chain reaction
RBD: Receptor-binding domain
RBM: Receptor-binding motif
RPA: Recombinase polymerase amplification
Th1: T helper type 1
TMA: Transcription–mediated amplification
TMPRSS2: Transmembrane protease, serine 2
SARS-CoV-2: Severe acute respiratory syndrome coronavirus 2
UTR: Untranslated region
WHO: World Health Organization
XUNG ĐỘT LỢI ÍCH
Các tác giả cam kết không có xung đột lợi ích.
ĐÓNG GÓP CỦA CÁC TÁC GIẢ
Tác giả Huỳnh Thị Ngọc Mai viết, tổng hợp và chỉnh sửa bản thảo.
Các tác giả Nguyễn Hoàng Thiên Phúc, Phan Hoàng Chí Hiếu, Phan Thị Hiếu Nghĩa, Lê Hồng Kông, Trương Thị Huỳnh Như tham gia viết bản thảo.
Các tác giả Khanh Lê, Hồ Văn Dũng tham gia chỉnh sửa bản thảo.
Các tác giả Nguyễn Thụy Vy, Trần Lê Bảo Hà, Trần Văn Hiếu, Nguyễn Hữu Hoàng tham gia viết và chỉnh sửa bản thảo.
Các tác giả Nguyễn Trí Nhân, Trần Linh Thước tham gia chỉnh sửa bản thảo.
References
- Huang C., Wang Y., Li X., Ren L., Zhao J., Hu Y.. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet. 2020;395(10223):497-506. Google Scholar
- Chan J.F.W., Yuan S.. A familial cluster of pneumonia associated with the 2019 novel coronavirus indicating person-to-person transmission: a study of a family cluster. Lancet. 2020;395(10223):514-523. Google Scholar
- Zhou P., Yang X.. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature. 2020;579(7798):270-273. PubMed Google Scholar
- Xu Y.. Unveiling the Origin and Transmission of 2019-nCoV. Vol. 28, Trends in Microbiology. Elsevier Ltd. 2020;:239-240. PubMed Google Scholar
- Coronavirus disease 2019 [Internet]. [cited 2020 Apr 15]. . ;:. Google Scholar
- Guan Y.. Isolation and characterization of viruses related to the SARS coronavirus from animals in Southern China. Science (80- ). 2003;302(5643):276-278. PubMed Google Scholar
- Cui J., Li F., Shi Z.L.. Origin and evolution of pathogenic coronaviruses. Vol. 17, Nature Reviews Microbiology. Nature Publishing Group. 2019;:181-192. PubMed Google Scholar
- Su S., Wong G., Shi W.. Epidemiology, Genetic Recombination, and Pathogenesis of Coronaviruses. Vol. 24, Trends in Microbiology. Elsevier Ltd. 2016;:490-502. PubMed Google Scholar
- Hu B., Ge X., Wang L.F., Shi Z.. Bat origin of human coronaviruses Coronaviruses: Emerging and re-emerging pathogens in humans and animals Susanna Lau Positive-strand RNA viruses. Vol. 12, Virology Journal. BioMed Central Ltd. 2015;1(10):. PubMed Google Scholar
- Berto A., Anh P.H.. Detection of potentially novel paramyxovirus and coronavirus viral RNA in bats and rats in the Mekong Delta region of southern Viet Nam. Zoonoses Public Health. ;65(1):30-42. PubMed Google Scholar
- Phan M.V.T., Ngo T.T., Hong A.P., Baker S., Kellam P., Cotten M.. Identification and characterization of Coronaviridae genomes from Vietnamese bats and rats based on conserved protein domains. Virus Evol. 2018;4(2):. PubMed Google Scholar
- COVID-19 Pandemic | Voice of America - English [Internet]. [cited 2020 Apr 10]. . ;:. Google Scholar
- COVID-19 Map - Johns Hopkins Coronavirus Resource Center [Internet]. [cited 2020 Jun 4]. . ;:. Google Scholar
- Realtime Monitoring Wuhan Coronavirus - By Kompa Group [Internet]. [cited 2020 Jun 3]. . ;:. Google Scholar
- TRANG TIN VỀ DỊCH BỆNH VIÊM ĐƯỜNG HÔ HẤP CẤP COVID-19 - Bộ Y tế [Internet]. [cited 2020 Jun 16] Available from:. . ;:. Google Scholar
- Kissler S.M., Tedijanto C., Goldstein E., Grad Y.H., Lipsitch M.. Projecting the transmission dynamics of SARS-CoV-2 through the post-pandemic period. medRxiv. 2020;:. Google Scholar
- Microdroplets might explain the rapid spread of COVID-19 | World Economic Forum [Internet]. [cited 2020 Apr 18]. . ;:. PubMed Google Scholar
- Hoffmann M., Kleine-Weber H., Schroeder S.. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell. 2020;:1-10. PubMed Google Scholar
- Walls A.C., Park Y.J., Tortorici M.A., Wall A., McGuire A.T., Veesler D.. Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell. 2020;:1-12. Google Scholar
- Wrapp D., Wang N.. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science (80- ). 2020;367(6483):1260-1263. PubMed Google Scholar
- Coutard B., Valle C., Lamballerie X., Canard B., Seidah N.G., Decroly E.. The spike glycoprotein of the new coronavirus 2019-nCoV contains a furin-like cleavage site absent in CoV of the same clade. Antiviral Res. 2020;176:104742. PubMed Google Scholar
- Tian X., Li C.. Potent binding of 2019 novel coronavirus spike protein by a SARS coronavirus-specific human monoclonal antibody. Emerg Microbes Infect. 2020;9(1):382-385. PubMed Google Scholar
- Lan J., Ge J.. Crystal structure of the 2019-nCoV spike receptor-binding domain bound with the ACE2 receptor. bioRxiv. 2020;:1-20. Google Scholar
- Ou X., Liu Y.. Characterization of spike glycoprotein of SARS-CoV-2 on virus entry and its immune cross-reactivity with SARS-CoV. Nat Commun. 2020;11(1):1620. PubMed Google Scholar
- Yan R., Zhang Y., Li Y., Xia L., Guo Y., Zhou Q.. Structural basis for the recognition of the SARS-CoV-2 by full-length human ACE2. Science. 2020;2762:1-10. PubMed Google Scholar
- Doremalen N., Bushmaker T., Morris D.H., Holbrook M.G., Gamble A., Williamson B.N.. Aerosol and Surface Stability of SARS-CoV-2 as Compared with SARS-CoV-1. N Engl J Med. 2020;:. Google Scholar
- Anfinrud P., Stadnytskyi V., Bax C.Ẻ., Bax A.. Visualizing Speech-Generated Oral Fluid Droplets with Laser Light Scattering. N Engl J Med. 2020;:. PubMed Google Scholar
- Ciotti M., Angeletti S.. COVID-19 Outbreak: An Overview. Chemotherapy. 2020;:1-9. PubMed Google Scholar
- Clinical Care Guidance for Healthcare Professionals about Coronavirus (COVID-19) | CDC [Internet]. [cited 2020 Jun 4]. . ;:. Google Scholar
- Management of Patients with Confirmed 2019-nCoV | CDC [Internet]. [cited 2020 Apr 15]. . ;:. PubMed Google Scholar
- Rabi F.A., Zoubi M.S., Al-Nasser A.D., Kasasbeh G.A., Salameh D.M.. Sars-cov-2 and coronavirus disease 2019: What we know so far. Pathogens. 2020;9(3):1-14. PubMed Google Scholar
- Morawska L., Cao J.. Airborne transmission of SARS-CoV-2: The world should face the reality. Environ Int. 2020;139:105730. PubMed Google Scholar
- Delamater P.L., Street E.J., Leslie T.F., Yang Y.T., Jacobsen K.H.. Complexity of the basic reproduction number (R0). Emerg Infect Dis. 2019;25(1):1-4. PubMed Google Scholar
- Zhao S., Lin Q.. Preliminary estimation of the basic reproduction number of novel coronavirus (2019-nCoV) in China, from 2019 to 2020: A data-driven analysis in the early phase of the outbreak. Int J Infect Dis. 2020;92:214-217. Google Scholar
- D'Arienzo M., Coniglio A.. Assessment of the SARS-CoV-2 basic reproduction number, R0, based on the early phase of COVID-19 outbreak in Italy. Biosaf Heal. 2020;:. Google Scholar
- Lauer S.A., Grantz K.H.. The Incubation Period of Coronavirus Disease 2019 (COVID-19) From Publicly Reported Confirmed Cases: Estimation and Application. Ann Intern Med. 2020;:. PubMed Google Scholar
- Team TNCPERE, Team TNCPERE. The Epidemiological Characteristics of an Outbreak of 2019 Novel Coronavirus Diseases (COVID-19) - China, 2020. China CDC Weekly. 2020;2(8):113-122. Google Scholar
- Onder G., Rezza G., Brusaferro S.. Case-Fatality Rate and Characteristics of Patients Dying in Relation to COVID-19 in Italy. Vol. 323, JAMA - Journal of the American Medical Association. American Medical Association. 2020;:1775-1776. PubMed Google Scholar
- Mortality Risk of COVID-19 - Statistics and Research - Our World in Data [Internet]. [cited 2020 Jun 6]. . ;:. Google Scholar
- Coronavirus Graphs: Worldwide Cases and Deaths - Worldometer [Internet]. [cited 2020 Jun 16]. . ;:. Google Scholar
- Coronavirus Death Rate (COVID-19) - Worldometer [Internet]. [cited 2020 Jun 6]. . ;:. Google Scholar
- Conti P., A. Younes. Coronavirus COV-19/SARS-CoV-2 affects women less than men: clinical response to viral infection. J Biol Regul Homeost Agents. 2020;34(2):. Google Scholar
- Mousavizadeh L., Ghasemi S.. Genotype and phenotype of COVID-19: Their roles in pathogenesis. J Microbiol Immunol Infect. 2020;:. PubMed Google Scholar
- Wu F., Zhao S.. A new coronavirus associated with human respiratory disease in China. Nature. 2020;579(7798):265-269. PubMed Google Scholar
- SARS-CoV-2 (Severe acute respiratory syndrome coronavirus 2) Sequences [Internet]. [cited 2020 Jun 3]. . ;:. Google Scholar
- GISAID - Next hCoV-19 App [Internet]. [cited 2020 Jun 3]. . ;:. Google Scholar
- Seah I., Su X., Lingam G.. Revisiting the dangers of the coronavirus in the ophthalmology practice. Eye. 2020;:1-3. Google Scholar
- Chan J.F.W., Kok K.H.. Genomic characterization of the 2019 novel human-pathogenic coronavirus isolated from a patient with atypical pneumonia after visiting Wuhan. Emerg Microbes Infect. 2020;9(1):221-236. PubMed Google Scholar
- Zeng Q., Langereis M.A., Van-Vliet A.L.W., Huizinga E.G., De-Groot R.J.. Structure of coronavirus hemagglutinin-esterase offers insight into corona and influenza virus evolution. Proc Natl Acad Sci U S A. 2008;105(26):9065-9069. PubMed Google Scholar
- Zhang T., Wu Q., Zhang Z.. Probable pangolin origin of 2019-nCoV associated with outbreak of COVID-19. SSRN eLibrary. 2020;:1-6. Google Scholar
- Zhang Y-Z., Holmes E.C.. A Genomic Perspective on the Origin and Emergence of SARS-CoV-2. Cell. 2020;0(0):1-5. Google Scholar
- Song W., Gui M., Wang X., Xiang Y.. Cryo-EM structure of the SARS coronavirus spike glycoprotein in complex with its host cell receptor ACE2. Heise MT, editor. PLOS Pathog. 2018;14(8):e1007236. PubMed Google Scholar
- Yuan Y., Cao D., Zhang Y.. Cryo-EM structures of MERS-CoV and SARS-CoV spike glycoproteins reveal the dynamic receptor binding domains. Nat Commun. 2017;8(1):1-9. PubMed Google Scholar
- Andersen K.G., Rambaut A., Lipkin W.I., Holmes E.C., Garry R.F.. The proximal origin of SARS-CoV-2. Nat Med. 2020;89(1):44-48. PubMed Google Scholar
- Zhou H., Chen X.. A Novel Bat Coronavirus Closely Related to SARS-CoV-2 Contains Natural Insertions at the S1/S2 Cleavage Site of the Spike Protein. Curr Biol. 2020;30(11):2196-2203. PubMed Google Scholar
- Jaimes J.A., Millet J.K., Whittaker G.R.. Proteolytic Cleavage of the SARS-CoV-2 Spike Protein and the Role of the Novel S1 / S2 Site Spike Protein and the Role of the Novel S1 / S2 Site. ISCIENCE. 2020;23(6):101212. PubMed Google Scholar
- Hemida M.G., Chu D.K.W.. Mers coronavirus in dromedary camel herd, Saudi Arabia. Emerg Infect Dis. 2014;20(7):1231-1234. PubMed Google Scholar
- Chu D.K.W., Poon L.L.M.. MERS coronaviruses in dromedary camels, Egypt. Emerg Infect Dis. 2014;20(6):1049-1053. PubMed Google Scholar
- Li F., Li W., M. Farzan, Harrison S.C.. Structural biology: Structure of SARS coronavirus spike receptor-binding domain complexed with receptor. Science (80- ). 2005;309(5742):1864-1868. PubMed Google Scholar
- Wu K., Peng G., Wilken M., Geraghty R.J., Li F.. Mechanisms of host receptor adaptation by severe acute respiratory syndrome coronavirus. J Biol Chem. 2012;287(12):8904-8911. PubMed Google Scholar
- Wan Y., Shang J., Graham R., Baric R.S., Li F.. Receptor recognition by novel coronavirus from Wuhan: An analysis based on decade-long structural studies of SARS. J Virol. 2020;:1-9. PubMed Google Scholar
- Diamond M.S., Pierson T.C.. The Challenges of Vaccine Development against a New Virus during a Pandemic. Cell Host Microbe. 2020;27(5):699-703. PubMed Google Scholar
- Duffy S.. Why are RNA virus mutation rates so damn high?. PLOS Biol. 2018;16(8):e3000003. PubMed Google Scholar
- Corman V.M., Landt O.. Detection of 2019 novel coronavirus (2019-nCoV) by real-time RT-PCR. Euro Surveill. 2020;25(3):. Google Scholar
- Suleman E., Mtshali M.S., Lane E.. Investigation of false positives associated with loop-mediated isothermal amplification assays for detection of Toxoplasma gondii in archived tissue samples of captive felids. J Vet Diagnostic Investig. 2016;28(5):536-542. PubMed Google Scholar
- Nguyen T., Duong B. D., Wolff A.. 2019 Novel Coronavirus Disease (COVID-19): Paving the Road for Rapid Detection and Point-of-Care Diagnostics. Micromachines. 2020;11(3):306. PubMed Google Scholar
- Hofmann W.P., Dries V., Herrmann E., Gärtner B., Zeuzem S., Sarrazin C.. Comparison of transcription mediated amplification (TMA) and reverse transcription polymerase chain reaction (RT-PCR) for detection of hepatitis C virus RNA in liver tissue. J Clin Virol. 2005;32(4):289-293. PubMed Google Scholar
- Miao F., Zhang J.. Rapid and sensitive recombinase polymerase amplification combined with lateral flow strip for detecting African swine fever virus. Front Microbiol. 2019;:. PubMed Google Scholar
- Comanor L., Anderson F.. Transcription-mediated amplification is more sensitive than conventional PCR-based assays for detecting residual serum HCV RNA at end of treatment. Am J Gastroenterol. 2001;96(10):2968-2972. PubMed Google Scholar
- Home - Vivaldi [Internet]. [cited 2020 Apr 13]. . ;:. PubMed Google Scholar
- Parida M.M., Santhosh S.R.. Development and evaluation of reverse transcription-loop-mediated isothermal amplification assay for rapid and real-time detection of Japanese encephalitis virus. J Clin Microbiol. 2006;;44(11):4172-4178. PubMed Google Scholar
- Zhou C., Mu Y.. Gold nanoparticles based colorimetric detection of target DNA after loop-mediated isothermal amplification. Chem Res Chinese Univ. 2013;29(3):424-428. Google Scholar
- Nishimasu H., Ran F.A.. Crystal structure of Cas9 in complex with guide RNA and target DNA. Cell. 2014;156(5):935-949. PubMed Google Scholar
- Jinek M., Jiang F., Taylor D.W.. Structures of Cas9 endonucleases reveal RNA-mediated conformational activation. Science (80- ). 2014;343(6176):. PubMed Google Scholar
- Wang X., Ji P.. CRISPR/Cas12a technology combined with immunochromatographic strips for portable detection of African swine fever virus. Commun Biol. ;3(1):1-8. PubMed Google Scholar
- Guo L., Sun X.. SARS-CoV-2 detection with CRISPR diagnostics. Cell Discov. 2020;6(1):34. PubMed Google Scholar
- Broughton J.P., Deng X.. CRISPR-Cas12-based detection of SARS-CoV-2. Nat Biotechnol. 2020;:. PubMed Google Scholar
- Aydin S.. A short history, principles, and types of ELISA, and our laboratory experience with peptide/protein analyses using ELISA. Peptides. 2015;72:4-15. PubMed Google Scholar
- Li M., Jin R.. Generation of antibodies against COVID-19 virus for development of diagnostic tools. medRxiv. 27;(1):2020.02.20.20025999. 2020;:. Google Scholar
- Diao B.. Diagnosis of Acute Respiratory Syndrome Coronavirus 2 Infection by Detection of Nucleocapsid Protein running title: Diagnosis of COVID-19 by N antigen detection. . ;:. Google Scholar
- Cai X., Chen J.. A Peptide-based Magnetic Chemiluminescence Enzyme Immunoassay for Serological Diagnosis of Corona Virus Disease 2019 (COVID-19). medRxiv. 2020.02.22.20026617.. 2020;:. Google Scholar
- Amanat F., Nguyen T.. A serological assay to detect SARS-CoV-2 seroconversion in humans. medRxiv. 2020.03.17.20037713.. 2020;:. Google Scholar
- Koczula K.M., Gallotta A.. Lateral flow assays. Essays Biochem. 2016;60(1):111-120. PubMed Google Scholar
- Xiang J., Yan M.. Evaluation of Enzyme-Linked Immunoassay and Colloidal Gold- Immunochromatographic Assay Kit for Detection of Novel Coronavirus (SARS-Cov-2) Causing an Outbreak of Pneumonia (COVID-19). medRxiv. 2020.02.27.20028787.. 2020;:. Google Scholar
- Alhassan A., Li Z., Poole C.B., Carlow C.K.S.. Expanding the MDx toolbox for filarial diagnosis and surveillance. Vol. 31, Trends in Parasitology. Elsevier Ltd. 2015;:391-400. PubMed Google Scholar
- Hull R.. Plant Virology: Fifth Edition. Plant Virology: Fifth Edition. Elsevier Inc. 2013;:797. Google Scholar
- Coronavirus Disease 2019 (COVID-19) | FDA [Internet]. [cited 2020 Apr 15]. . ;:. Google Scholar
- Clinical management of severe acute respiratory infection when COVID-19 is suspected [Internet]. [cited 2020 Apr 15]. . ;:. Google Scholar
- Pang J., Wang M.X.. Potential Rapid Diagnostics, Vaccine and Therapeutics for 2019 Novel Coronavirus (2019-nCoV): A Systematic Review. J Clin Med. 2020;9(3):623. PubMed Google Scholar
- Liu C., Zhou Q.. Research and Development on Therapeutic Agents and Vaccines for COVID-19 and Related Human Coronavirus Diseases. ACS Cent Sci. 2020;:. PubMed Google Scholar
- Wu R., Wang L.. An Update on Current Therapeutic Drugs Treating COVID-19. Curr Pharmacol reports. 2020;:1-15. PubMed Google Scholar
- Sheahan T.P., Sims A.C.. Comparative therapeutic efficacy of remdesivir and combination lopinavir, ritonavir, and interferon beta against MERS-CoV. Nat Commun. 2020;11(1):222. PubMed Google Scholar
- Wit E., Feldmann F.. Prophylactic and therapeutic remdesivir (GS-5734) treatment in the rhesus macaque model of MERS-CoV infection. Proc Natl Acad Sci U S A. 2020;117(12):6771-6776. PubMed Google Scholar
- Ko W.C., Rolain J.M.. Arguments in favour of remdesivir for treating SARS-CoV-2 infections. International Journal of Antimicrobial Agents. Elsevier B.V. 2020;:. PubMed Google Scholar
- NIH clinical trial shows Remdesivir accelerates recovery from advanced COVID-19 | National Institutes of Health (NIH) [Internet]. [cited 2020 May 21]. . ;:. Google Scholar
- Vincent M.J., Bergeron E.. Chloroquine is a potent inhibitor of SARS coronavirus infection and spread. Virol J. Aug. 2005;22(2):69. PubMed Google Scholar
- Keyaerts E., Vijgen L., Maes P., Neyts J., Ranst M.V.. In vitro inhibition of severe acute respiratory syndrome coronavirus by chloroquine. Biochem Biophys Res Commun. 2004;323(1):264-268. PubMed Google Scholar
- Şimşek-Yavuz S., Ünal S.. Antiviral treatment of covid-19. Vol. 50, Turkish Journal of Medical Sciences. Turkiye Klinikleri. 2020;:611-619. PubMed Google Scholar
- The Vietnam Chloroquine Treatment on COVID-19 - Full Text View - ClinicalTrials.gov [Internet]. [cited 2020 Jun 5]. . ;:. Google Scholar
- Gbinigie K., Frie K.. Should chloroquine and hydroxychloroquine be used to treat COVID-19? A rapid review. BJGP open. 2020;:. PubMed Google Scholar
- Cao B., Wang Y.. A Trial of Lopinavir-Ritonavir in Adults Hospitalized with Severe Covid-19. N Engl J Med. 2020;:. Google Scholar
- Lim J., Jeon S.. Case of the index patient who caused tertiary transmission of coronavirus disease 2019 in Korea: The application of lopinavir/ritonavir for the treatment of COVID-19 pneumonia monitored by quantitative RT-PCR. J Korean Med Sci. ;35(6):. PubMed Google Scholar
- Cai Q., Yang M.. Experimental Treatment with Favipiravir for COVID-19: An Open-Label Control Study. Engineering. 2020;:. PubMed Google Scholar
- Li Y., Liu X.. Traditional Chinese herbal medicine for treating novel coronavirus (COVID-19) pneumonia: protocol for a systematic review and meta-analysis. Syst Rev. Dec. 2020;9(1):75. PubMed Google Scholar
- Xu J., Zhang Y.. Traditional Chinese Medicine treatment of COVID-19. . Complement Ther Clin Pract. 2020;39:101165. PubMed Google Scholar
- Ho L.T.F., Chan K.K.H., Chung V.C.H., Leung T.H.. Highlights of traditional Chinese medicine frontline expert advice in the China national guideline for COVID-19. Eur J Integr Med. 2020;3:101116. PubMed Google Scholar
- Luo H.. Can Chinese Medicine Be Used for Prevention of Corona Virus Disease 2019 (COVID-19)? A Review of Historical Classics, Research Evidence and Current Prevention Programs. Chin J Integr Med. 2020;:. PubMed Google Scholar
- Thuy B.T.P., My T.T.A.. Investigation into SARS-CoV-2 Resistance of Compounds in Garlic Essential Oil. ACS Omega. 2020;:. Google Scholar
- Therapeutic Options for COVID-19 Patients | CDC [Internet]. [cited 2020 Jun 5]. . ;:. Google Scholar
- Singhal T.. A Review of Coronavirus Disease-2019 (COVID-19). Vol. 87, Indian Journal of Pediatrics. Springer. 2020;:281-286. PubMed Google Scholar
- Wujtewicz M., Dylczyk-Sommer A., Aszkiełowicz A., Zdanowski S., Piwowarczyk S., Owczuk R.. COVID-19 - what should anaethesiologists and intensivists know about it?. Anaesthesiol Intensive Ther. 2020;52(1):34-41. PubMed Google Scholar
- Song Z., Hu Y., Zheng S., Yang L., Zhao R.. Hospital pharmacists' pharmaceutical care for hospitalized patients with COVID-19: Recommendations and guidance from clinical experience. Res Soc Adm Pharm. 2020;:. PubMed Google Scholar
- Jin YH, Cai L, Cheng ZS, Cheng H, Deng T, Fan YP, et al. A rapid advice guideline for the diagnosis and treatment of 2019 novel coronavirus (2019-nCoV) infected pneumonia (standard version). Vol. 7, Military Medical Research. BioMed Central Ltd. 2020;:4. Google Scholar
- Shanmugaraj B., Siriwattananon K.. Perspectives on monoclonal antibody therapy as potential therapeutic intervention for Coronavirus disease-19 (COVID-19). Vol. 38, Asian Pacific journal of allergy and immunology. NLM (Medline). 2020;:10-18. Google Scholar
- AminJafari A., Ghasemi S.. The Possible of Immunotherapy for COVID-19: a Systematic Review. Int Immunopharmacol. 2020;83:106455. PubMed Google Scholar
- Bloch E.M., Shoham S.. Deployment of convalescent plasma for the prevention and treatment of COVID-19. J Clin Invest. 2020;:. PubMed Google Scholar
- Duan K., Liu B., Li C.. Effectiveness of convalescent plasma therapy in severe COVID-19 patients. Proc Natl Acad Sci. 202004168. 2020;:. Google Scholar
- Zhang B., Liu S.. Treatment with convalescent plasma for critically ill patients with SARS-CoV-2 infection. Chest. 2020;:. PubMed Google Scholar
- Yi Y., Lagniton P.N.P., Ye S., Li E., Xu R.H.. COVID-19: what has been learned and to be learned about the novel coronavirus disease. Int J Biol Sci. 2020;16(10):1753-1766. PubMed Google Scholar
- Strugnell R., Zepp F., Cunningham A., Tantawichien T.. Vaccine antigens. Perspect Vaccinol. 2011;1(1):61-88. Google Scholar
- WHO | Severe Acute Respiratory Syndrome (SARS) [Internet]. [cited 2020 Apr 15]. . ;:. Google Scholar
- WHO | Middle East respiratory syndrome coronavirus (MERS-CoV) [Internet]. [cited 2020 Apr 15]. . ;:. Google Scholar
- Draft landscape of COVID-19 candidate vaccines [Internet]. [cited 2020 Jun 3]. . ;:. Google Scholar
- He Y., Jiang S.. Vaccine Design for Severe Acute Respiratory Syndrome Coronavirus. . 2005;18(2):327-332. PubMed Google Scholar
- Chinese Clinical Trial Register (ChiCTR) - The world health organization international clinical trials registered organization registered platform [Internet]. [cited 2020 Jun 3]. . ;:. Google Scholar
- . . ;:. Google Scholar
- Safety and Immunogenicity Study of Inactivated Vaccine for Prophylaxis of SARS CoV-2 Infection (COVID-19) - Full Text View - ClinicalTrials.gov [Internet]. [cited 2020 Jun 3]. . ;:. Google Scholar
- Safety and Immunogenicity Study of Inactivated Vaccine for Prevention of SARS-CoV-2 Infection(COVID-19) - Full Text View - ClinicalTrials.gov [Internet]. [cited 2020 Jun 3]. . ;:. Google Scholar
- Gao Q., Bao L.. Development of an inactivated vaccine candidate for SARS-CoV-2. Science (80- ). 2020;eabc1932:. PubMed Google Scholar
- He Y., Zhou Y., Siddiqui P., Jiang S.. Inactivated SARS-CoV vaccine elicits high titers of spike protein-specific antibodies that block receptor binding and virus entry. Biochem Biophys Res Commun. 2004;325(2):445-452. PubMed Google Scholar
- Tang L., Zhu Q.. Inactivated SARS-CoV vaccine prepared from whole virus induces a high level of neutralizing antibodies in BALB/c mice. DNA Cell Biol. 2004;23(6):391-394. PubMed Google Scholar
- Tsunetsugu-Yokota Y.. Large-Scale Preparation of UV-Inactivated SARS Coronavirus Virions for Vaccine Antigen. In: Cavanagh D, editor. SARS- and Other Coronaviruses: Laboratory Protocols. Totowa, NJ: Humana Press. 2008;:119-126. PubMed Google Scholar
- Xiong S., Wang Y.F.. Immunogenicity of SARS inactivated vaccine in BALB/c mice. Immunol Lett. 2004;95(2):139-143. PubMed Google Scholar
- He Y., Li J.. Identification and characterization of novel neutralizing epitopes in the receptor-binding domain of SARS-CoV spike protein: Revealing the critical antigenic determinants in inactivated SARS-CoV vaccine. Vaccine. 2006. ;24(26):5498-5508. PubMed Google Scholar
- Jiang S., He Y., Liu S.. SARS Vaccine Development - Volume 11, Number 7-July 2005 - Emerging Infectious Diseases journal - CDC. Emerg Infect Dis. 2005;11(7):1016. PubMed Google Scholar
- Shang W., Yang Y., Rao Y., Rao X.. The outbreak of SARS-CoV-2 pneumonia calls for viral vaccines. npj Vaccines. 2020;5(1):18. PubMed Google Scholar
- Kurup D., Wirblich C., Feldmann H., Marzi A., Schnell M.J.. Rhabdovirus-Based Vaccine Platforms against Henipaviruses. J Virol. 2015;89(1):144-154. PubMed Google Scholar
- Tatsis N., Ertl H.C.J.. Adenoviruses as vaccine vectors. Vol. 10, Molecular Therapy. Cell Press. 2004;:616-629. PubMed Google Scholar
- Baden L.R., Walsh S.R.. First-in-Human Evaluation of the Safety and Immunogenicity of a Recombinant Adenovirus Serotype 26 HIV-1 Env Vaccine (IPCAVD 001). . 2012;:. Google Scholar
- Buchbinder S.P., Mehrotra D.V.. Efficacy assessment of a cell-mediated immunity HIV-1 vaccine (the Step Study): a double-blind, randomised, placebo-controlled, test-of-concept trial. Lancet. 2008;372(9653):1881-1893. Google Scholar
- Callebaut P., Enjuanes L., Pensaert M.. An adenovirus recombinant expressing the spike glycoprotein of porcine respiratory coronavirus is immunogenic in swine. J Gen Virol. 1996;77(2):309-313. PubMed Google Scholar
- Weingartl H., Czub M.. Immunization with Modified Vaccinia Virus Ankara-Based Recombinant Vaccine against Severe Acute Respiratory Syndrome Is Associated with Enhanced Hepatitis in Ferrets. J Virol. 2004;78(22):12672-12676. PubMed Google Scholar
- Clinical Trials Register ChAdOx1 [Internet]. [cited 2020 Jun 3]. . ;:. Google Scholar
- Zhu F.C., Li Y.H.. Safety, tolerability, and immunogenicity of a recombinant adenovirus type-5 vectored COVID-19 vaccine: a dose-escalation, open-label, non-randomised, first-in-human trial. Lancet. 2020;:. Google Scholar
- Chinese Clinical Trial Register (ChiCTR) - The world health organization international clinical trials registered organization registered platform [Internet]. [cited 2020 Jun 3]. . ;:. Google Scholar
- Buchholz U.J., Bukreyev A., Yang L., Lamirande E.W., Murphy B.R.. Contributions of the structural proteins of severe respiratory syndrome coronavirus to protective immunity. Proc Natl Acad Sci U S A. 2004;101(26):9804-9809. PubMed Google Scholar
- Li H.Y., Ramalingam S., Chye M.L.. Accumulation of recombinant SARS-CoV spike protein in plant cytosol and chloroplasts indicate potential for development of plant-derived oral vaccines. Exp Biol Med. 2006;231(8):1346-1352. PubMed Google Scholar
- Pogrebnyak N., Golovkin M.. Severe acute respiratory syndrome (SARS) S protein production in plants: Development of recombinant vaccine. Proc Natl Acad Sci U S A. 2005;102(25):9062-9067. PubMed Google Scholar
- Twyman R., Schillberg S., Fischer R.. Optimizing the Yield of Recombinant Pharmaceutical Proteins in Plants. Curr Pharm Des. 2013;19(31):5486-5494. PubMed Google Scholar
- Evaluation of the Safety and Immunogenicity of a SARS-CoV-2 rS (COVID-19) Nanoparticle Vaccine With/Without Matrix-M Adjuvant - Full Text View - ClinicalTrials.gov [Internet]. [cited 2020 Jun 3]. . ;:. Google Scholar
- Smith T.R.F.. Immunogenicity of a DNA vaccine candidate for COVID-19. Nat Commun. ;11(1):2601. PubMed Google Scholar
- Martin J.E.. A SARS DNA Vaccine Induces Neutralizing Antibody and Cellular Immune Responses in Healthy Adults in a Phase I Clinical Trial. . 2009;26(50):6338-6343. PubMed Google Scholar
- King D.F.L., McKay P.F., Mann J.F.S., Jones C.B., Shattock R.J.. Plasmid DNA Vaccine Co-Immunisation Modulates Cellular and Humoral Immune Responses Induced by Intranasal Inoculation in Mice. Wang S, editor. PLoS One. 2015;10(11):e0141557. PubMed Google Scholar
- Yang Z.Y.. A DNA vaccine induces SARS coronavirus neutralization and protective immunity in mice. Nature. 2004;428(6982):561-564. PubMed Google Scholar
- Marshall E., Enserink M.. Caution Urged on SARS Vaccines. Science (80- ). 2004;303(5660):944-946. PubMed Google Scholar
- Safety, Tolerability and Immunogenicity of INO-4800 for COVID-19 in Healthy Volunteers - Full Text View - ClinicalTrials.gov [Internet]. [cited 2020 Jun 3]. . ;:. Google Scholar
- Martinon F.. Induction of virus‐specific cytotoxic T lymphocytes in vivo by liposome‐entrapped mRNA. Eur J Immunol. 1993;23(7):1719-1722. PubMed Google Scholar
- Pardi N.. Zika virus protection by a single low-dose nucleoside-modified mRNA vaccination. Nature. 2017;543(7644):248-251. Google Scholar
- Alberer M.. Safety and immunogenicity of a mRNA rabies vaccine in healthy adults: an open-label, non-randomised, prospective, first-in-human phase 1 clinical trial. Lancet. 2017;390(10101):1511-1520. Google Scholar
- John S.. Multi-antigenic human cytomegalovirus mRNA vaccines that elicit potent humoral and cell-mediated immunity. Vaccine. 2018;36(12):1689-1699. PubMed Google Scholar
- Tsai H.Ẻ.. Pro-opiomelanocortin gene delivery suppresses the growth of established Lewis lung carcinoma through a melanocortin-1 receptor-independent pathway. J Gene Med. 2012;14(1):44-53. PubMed Google Scholar
- J. Probst. Spontaneous cellular uptake of exogenous messenger RNA in vivo is nucleic acid-specific, saturable and ion dependent. Gene Ther. 2007;14(15):1175-1180. PubMed Google Scholar
- Petsch B.. Protective efficacy of in vitro synthesized, specific mRNA vaccines against influenza A virus infection. Nat Biotechnol. 2012;30(12):1210-1216. PubMed Google Scholar
- Banerjee A.K.. 5'-Terminal cap structure in eucaryotic messenger ribonucleic acids. Microbiological Reviews. 1980;44:175-205. PubMed Google Scholar
- Wickens M.. How the messenger got its tail: addition of poly(A) in the nucleus. Trends Biochem Sci. 1990;15(7):277-281. Google Scholar
- Gallie D.R.. The cap and poly(A) tail function synergistically to regulate mRNA translational efficiency. Genes Dev. 1991;5(11):2108-2116. PubMed Google Scholar
- Kozak M.. Point mutations define a sequence flanking the AUG initiator codon that modulates translation by eukaryotic ribosomes. Cell. 1986;44(2):283-292. Google Scholar
- Vivinus S., Baulande S.. An element within the 5′ untranslated region of human Hsp70 mRNA which acts as a general enhancer of mRNA translation. Eur J Biochem. 2001;268(7):1908-1917. PubMed Google Scholar
- Zhang H.. Genome editing of upstream open reading frames enables translational control in plants. Vol. 36, Nature Biotechnology. Nature Publishing Group. 2018;:894-900. PubMed Google Scholar
- Karikó K., Muramatsu H., Ludwig J., Weissman D.. Generating the optimal mRNA for therapy: HPLC purification eliminates immune activation and improves translation of nucleoside-modified, protein-encoding mRNA. Nucleic Acids Res. 2011;39(21):1-10. PubMed Google Scholar
- Wang Y.. Systemic delivery of modified mRNA encoding herpes simplex virus 1 thymidine kinase for targeted cancer gene therapy. Mol Ther. 2013;21(2):358-367. PubMed Google Scholar





