
 Open Access
Open Access Abstract
Keratinocyte Growth Factor (KGF) is a paracrine-acting and epithelium-specific growth factor produced by cells of mesenchymal origin, play an important role in promoting proliferation, differentiation, motility of epithelial cells and stimulating regeneration of damaged epithelial tissues. Recent studies indicated that recombinant KGF is produced in many different expression systems such as bacteria, insect cells, plant and mammalian cells. However, KGF’s yields obtained from these systems is low and production’s cost is high especially in mammalian cells. In this study, the yeast Pichia pastoris was chosen as a host for KGF expression through induction of methanol by promoter AOX on pPICzαA vector system. The results demonstrated that the Pichia pastoris X33:kgf transformants secreted KGF directly into BMMY medium after inducing by 0.5% methanol. The recombinant protein was purified by heparin affinity chromatography with the yield of 1.35 mg/l and the purity of 99.89% showed by SDS-PAGE. In addition, MTT assay showed the purified recombinant KGF had a proliferation effect on A549 cell line since A549 known as a cell has KGF’s receptor.
GIỚI THIỆU
KGF hay FGF7, là nhân tố tăng trưởng được sinh tổng hợp bởi các tế bào có nguồn gốc trung mô, được phát hiện và phân lập lần đầu tiên vào năm 1989 bởi Rubin và cộng sự từ dòng tế bào nguyên bào sợi phổi M426 trong phôi thai người 1 , 2 . Các nghiên cứu cho thấy rằng KGF có nhiều chức năng khác nhau bao gồm kích thích phân chia, tăng sinh và biệt hóa các dòng tế bào biểu mô và tế bào sừng 1 , 3 , 4 . Bên cạnh đó, KGF còn có vai trò trong việc tái tạo, sửa chữa các mô và cơ quan tổn thương thông kích thích sự di chuyển của các tế bào sừng và biểu mô ở miệng vết thương, từ đó thúc đẩy tiến trình làm lành vết thương 5 , 6 . Hiện nay, KGF đã được nghiên cứu rộng rãi và ứng dụng trong nhiều lĩnh vực như mỹ phẩm, dược phẩm và hỗ trợ trong điều trị ung thư 7 , 8 , 9 .
Việc hóa trị và xạ trị thời gian dài trong điều trị ung thư có thể gây ra nhiều tác dụng phụ đối với cơ thể bệnh nhân mà phổ biến nhất là giảm sức đề kháng, dẫn đến người bệnh phải đối mặt thêm với các hội chứng khác như loét miệng, nhiễm trùng hay rụng tóc 10 . Để giải quyết tình trạng trên, thị trường xuất hiện nhiều sản phẩm hướng đến hỗ trợ bệnh nhân trong điều trị ung thư, trong đó có nhiều sản phẩm có bản chất sinh học. Palifermin (KepivanceTM) là sản phẩm KGF tái tổ hợp sản xuất trên hệ thống E. coli đã được FDA Hoa Kì phê chuẩn vào ngày 15/12/2004, chấp thuận cho thương mại và sử dụng để làm giảm hội chứng viêm loét màng nhầy niêm mạc miệng ở các bệnh nhân ung thư máu ác tính trải qua nhiều trị lần hóa trị và xạ trị.
Với tiềm năng ứng dụng lớn trong nhiều lĩnh vực, tuy nhiên việc sử dụng KGF vẫn còn hạn chế vì giá thành cao do lượng KGF thu nhận tự nhiên tương đối thấp. Theo theo thống kê, trung bình một bệnh nhân có trọng lượng 70 kg sẽ tốn khoảng 5000 euro cho một lần điều trị bằng Palifermin 11 , tương đương khoảng hơn 100 triệu VNĐ. Với mục tiêu thu nhận KGF với lượng lớn hơn, có nhiều chiến lược biểu hiện tái tổ hợp đã và đang được tiến hành trên nhiều hệ thống như vi khuẩn E. c oli 12 , 13 , tế bào cây thuốc lá 6 , tế bào con tằm 14 , tế bào chuột lang Trung Quốc – CHO 15 , tuy nhiên vẫn còn tồn tại nhiều nhược điểm như lượng KGF thu nhận thấp, bản thân KGF gây độc cho tế bào chủ biểu hiện như ở hệ thống E. coli 12 ; đồng thời, hiệu quả sản xuất thấp do chi phí cao ở hệ thống biểu hiện tế bào động vật có vú,…
Năm 2018, Bahadori và cộng sự đã biểu hiện thành công KGF dạng tiết ở hệ thống tế bào nấm men P. pastoris với hoạt tính kích thích tăng sinh trên dòng tế bào A549 và NIH3T3. Nhận thấy P. pastoris là hệ thống tế bào chủ biểu hiện với nhiều ưu điểm như khả năng biến đổi sau dịch mã và gấp cuộn protein tương đồng với tế bào động vật, đặc biệt promoter mạnh và khả năng tiết protein ngoại bào giúp protein tái tổ hợp ở hệ thống này biểu hiện ở mức độ cao, có thể chiếm tới hơn 80% tổng protein tiết 16 , 17 , 18 , chúng tôi lựa chọn P. pastoris nhằm biểu hiện KGF tái tổ hợp. Với chiến lược trên, gen mã hóa cho protein KGF được dòng hóa vào vector pPICZαA sau đó gắn chèn vào DNA bộ gen của nấm men P. pastoris X33 nhờ cơ chế tái tổ hợp tương đồng. Dưới sự kiểm soát của promoter AOX1- cảm ứng bởi methanol, protein KGF dạng tiết được biểu hiện, thu nhận và tinh sạch bằng sắc ký ái lực heparin, sau đó, đánh giá hoạt tính sinh học dựa trên khả năng kích thích tăng sinh tế bào A549. Kết quả của đề tài hướng tới giảm giá thành của các sản phẩm thương mại hiện tại và góp phần hỗ trợ các bệnh nhân trong việc điều trị ung thư cũng như mở rộng ứng dụng trong các lĩnh vực khác như mỹ phẩm hay các sản phẩm chăm sóc sức khỏe.
VẬT LIỆU – PHƯƠNG PHÁP
Chủng, plasmid và tế bào
Chủng E.coli DH5α [F – φ80 lac Z ΔM15 Δ( lac ZYA- arg F) U169 rec A1 end A1 hsd R17 (r K – , m K + ) pho A sup E44 λ – thi -1 gyr A96 rel A1] dùng để cấu trúc vector tái tổ hợp, được nuôi cấy trong môi trường LB (Trypton 1%, cao nấm men 0,5%, NaCl 0,5%).
Chủng nấm men P. pastoris X33 [Hoang dại, Mut + ] dùng để biểu hiện KGF, được nuôi cấy trong môi trường YPD (Dextrose 2%, peptone 2%, cao nấm men 1%).
Plasmid pPICZαA (V195-20, Invitrogen) có kích thước 3593bp được dùng làm vector dòng hóa gen kgf , mang gen kháng zeocin hỗ trợ quá trình sàng lọc dòng tái tổ hợp, promoter AOX cảm ứng bởi methanol, trình tự tín hiệu tiết α-MF giúp hỗ trợ tiết protein ngoại bào.
Gen mã hóa cho protein KGF (gen kgf) được tham khảo từ Genbank [Genbank: M60828.1], thiết kế lại dựa trên trình tự protein KGF từ ngân hàng dữ liệu protein Drugbank [Drugbank: Palifermin DB00039] và tối ưu hóa nhằm phù hợp với bộ mã di truyền của chủng chủ biểu hiện P . p astoris .
Dòng tế bào A549 (ATCC: CCL-1859) có thụ thể KGFR được sử dụng để thử hoạt tính sinh học của protein KGF.
Cấu trúc vector tái tổ hợp pPICZαA-kgf
Sau khi được tổng hợp hóa học từ trình tự đã thiết kế, gen kgf được khuếch đại bằng phương pháp PCR bằng cặp mồi đặc hiệu KGF-F/R với trình tự lần lượt là AAG GGG TAT CTC TCG AGA AAA GAT CTT ACG ATT ACA TGG AAGG và GCG GCC GCC GCG GCT CGA GTC ATT AAG TAA TAG CCA TTG GCAA. Chu kỳ PCR được thiết lập như sau: Biến tính 94 o C 3 phút, lặp lại 30 lần chu kỳ 94 o C 30 giây, 55 o C 30 giây, 72 o C 30 giây, kéo dài 72 o C 5 phút, ổn định 32 o C 3 phút. Vector pPICZαA được tách chiết bằng phương pháp SDS-kiềm và được cắt mở vòng bằng enzyme Xho I. Sản phẩm PCR và sản phẩm cắt được kiểm tra bằng phương pháp điện di trên gel agarose 1,2%.
Gen kgf được tinh sạch bằng phương pháp tủa cồn, plasmid sau khi cắt được tinh sạch qua cột silica gel bằng bộ kit EZ-10 (Biobasic), sau đó gen kgf được dòng hóa vào plasmid pPICZαA theo cơ chế tái tổ hợp tương đồng bằng bộ kit eClone được cung cấp bởi bộ môn Công nghệ Sinh học Phân từ và Môi trường – trường Đại học Khoa học Tự nhiên, ĐHQG TP. HCM. Cấu trúc vector tái tổ hợp và sơ đồ tạo dòng được thể hiện ở Figure 1 . Sản phẩm nối được biến nạp vào tế bào E.coli DH5α bằng phương pháp hóa biến nạp và sàng lọc trên môi trường LB agar với kháng sinh zeocin (nồng độ 50 µg/ml). Các thể biến nạp được sàng lọc bước đầu bằng phản ứng PCR khuẩn lạc bằng mồi đặc hiệu, các khuẩn lạc cho kết quả dương tính được tiến hành tách chiết plasmid bằng phương pháp SDS – kiềm. Các plasmid sau khi tách chiết được sử dụng làm mạch khuôn cho phản ứng PCR với cặp mồi KGF-F và AOX-R để xác nhận gen đã chèn vào plasmid pPICZαA đúng khung đọc mở như thiết kế. Cuối cùng, vector tái tổ hợp được giải trình tự bằng cặp mồi AOX-F và AOX-R.
Figure 1 . Cấu trúc vector tái tổ hợp pPICzαA/kgf và sơ đồ tạo dòng E.coli DH5α mang vector tái tổ hợp

Tạo dòng nấm men P. pastoris X33 mang vector tái tổ hợp pPICz αA-kgf
Để cấu trúc chủng nấm men P. pastoris :: kgf , các plasmid tái tổ hợp mang gen kgf được cắt mở vòng với enzyme Sac I tại vị trí vùng promoter AOX1 nhằm tăng khả năng sát nhập vào bộ gen của nấm men, sau đó được đưa vào tế bào chủ P. pastoris bằng phương pháp điện biến nạp với điện dung 25 μF, điện trở 200 Ω, hiệu điện thế 1,5 kV trong 5,0 ms.
Nhằm phát hiện sự hiện diện của gen trong các thể biến nạp, các khuẩn lạc mọc được trên đĩa YPD với kháng sinh zeocin với nồng độ 100 μg/ml được kiểm tra bằng kĩ thuật PCR khuẩn lạc sàng lọc với mồi đặc hiệu KGF-F và KGF-R. Các khuẩn lạc nấm men được phá lớp vách dày chứa chitin và đường mannose bằng lyticase trước khi sử dụng làm khuôn cho phản ứng PCR. Các khuẩn lạc cho kết quả PCR dương tính được tách chiết DNA bộ gen và kiểm tra kiểu hình bằng phản ứng PCR với cặp mồi AOX-F/R.
Kiểm tra biểu hiện KGF dạng tiết của chủng P. pastoris X33::kgf
Các dòng P. pastoris X33:: kgf tái tổ hợp mang kiểu hình Mut+ được tiến hành hoạt hóa trong môi trường BMGY (cao nấm men 1%, peptone 2%, YNB 1,34%, phosphate buffer 0.1M pH 6,0; biotin 4×10 -5 %, glycerol 1%), nuôi cấy lắc trong 16 – 18 giờ ở nhiệt độ 30 o C, tốc độ 250 vòng/phút sau đó được cấy chuyền sang môi trường BMMY (cao nấm men 1%, peptone 2%, YNB 1,34%, 10% phosphate buffer 1 M pH 6,0; biotin 4×10 -5 %, methanol 0,5%) với mật độ tế bào khởi điểm OD 600 =1 và bổ sung methanol với nồng độ cuối 0,5% sau mỗi 24 giờ nuôi cấy. Mật độ tế bào được xác định sau lần lượt 24 giờ, 48 giờ và 72 giờ nhằm kiểm soát quá trình tăng trưởng của dòng nấm men tái tổ hợp, bên cạnh đó, mức độ biểu hiện của protein KGF trong dịch biểu hiện cũng được đánh giá bằng phương pháp di SDS-PAGE và khẳng định lại bằng phương pháp lai western với kháng thể đặc hiệu kháng KGF. Cụ thể, mẫu protein được phân tách trên gelpolyacrylamide và được chuyển thẩm lên màng nitrocellulose, khóa màng trong sữa gầy 5% qua đêm, sau đó tiến hành lai với kháng thể sơ cấp kháng KGF (R&D Systems) nồng độ 1µg/ml và kháng thể thứ cấp mouse – IgG gắn Horseradish peroxidase (HRP) (R&D Systems) tỉ lệ 1/1000. Tín hiệu được ghi nhận trên màng lai nhờ phản ứng giữa cơ chất peroxidase (H 2 O 2 ) và luminol với HRP.
Tinh chế thu nhận protein KGF tái tổ hợp bằng sắc ký ái lực Heparin
Cột sắc ký ái lực Heparin 5ml (GE Healthcare Life Sciences) được cân bằng với 50 ml dung dịch A (20mM Tris – HCl, pH 7,5), 200 ml dịch biểu hiện sau khi thu nhận sẽ được lọc qua màng lọc 0,2µm sau đó tiến hành nạp qua cột. Sau khi toàn bộ mẫu đã được nạp, tiến hành rửa cột với 50ml 100% dung dịch A và 50 ml dung dịch 30% B (20mM Tris – HCl, pH 7,5, 2M NaCl) nhằm loại bỏ một phần protein tạp bám không đặc hiệu trên cột, sau đó protein mục tiêu KGF được dung ly khỏi cột với 50% và 100% dung dịch B. Tốc độ cho toàn quy trình là 5ml/phút. Các phân đoạn sau tinh chế được tiến hành điện di SDS PAGE, đánh giá độ tinh sạch bằng phần mềm Gel analyzer và định lượng nồng độ protein mục tiêu bằng phương pháp Bradford.
Thử nghiệm hoạt tính sinh học của protein KGF tái tổ hợp bằng phương pháp MTT
Thử nghiệm MTT được dùng để đánh giá hoạt tính kích thích tăng sinh của KGF tái tổ hợp trên tế bào A549. Tế bào A549 được nuôi cấy 3 ngày trong đĩa Φ60, thu nhận và chuyển vào vào đĩa 96 giếng với mật độ 10 4 tế bào/giếng trong môi trường nuôi cấy DMEM/F12 có bổ sung 10% FBS, 100μl/ giếng. Ủ tế bào ở 37 o C, 5% CO 2 trong 24 giờ. Loại môi trường cũ, rửa tế bào bằng môi trường DMEM/F12 không chứa FBS, bổ sung 95 μl DMEM/F12 không chứa FBS. Bổ sung 5μl KGF ở các nồng độ 25; 50; 100; 200 ng/μl vào mỗi giếng và 5μl dung dịch pha loãng mẫu vào chứng âm. Ủ tế bào ở 37 o C, 5% CO 2 trong 48 giờ. Bổ sung 5μl MTT (5mg/ml) vào mỗi giếng. Ủ ở 37°C, 5% CO 2 trong 3 giờ. Hút bỏ toàn bộ dịch nổi, bổ sung 100μl dung dịch hoà tan MTT vào mỗi giếng. Đặt đĩa lên máy lắc trong 10 phút. Đo lượng tinh thể tím được tạo thành bằng máy đọc đĩa 96 giếng ở bước sóng 550 nm. Thực hiện tương tự với mẫu KGF thương mại. Mỗi thí nghiệm được lặp lại 3 lần và đánh giá thống kê bằng T-test.
KẾT QUẢ VÀ THẢO LUẬN
Kết quả tối ưu hóa trình tự gen mã hóa cho phân đoạn protein KGF
Trình tự gen kgf được dịch mã ngược từ trình tự polypeptide của KGF thu từ ngân hàng dữ liệu Drugbank. Đây là trình tự protein KGF được biểu hiện và thu nhận ở hệ thống E.coli và được sử dụng làm thuốc. Vì vậy với mục tiêu biểu hiện protein KGF với cùng trình tự polypeptide ở hệ thống Pichia pastoris , chúng tôi tiến hành tối ưu một số bộ ba mã hóa của gene kgf . Gen mục tiêu được tối ưu bằng phần mềm snapgene và kiểm tra trình tự bằng phần mềm Genscript. Kết quả phân tích trình tự ở Table 1 cho thấy, trước khi được tối ưu, gen kgf có chỉ số tương thích codon (CAI) trên hệ thống Pichia pastoris đạt 0,8 – là chỉ số thấp nhất trong khoảng lý tưởng (0,8 – 1,0). Sau khi được tối ưu, chỉ số tương thích đạt 0,92, bên cạnh đó tần suất sử dụng codon hiếm cũng giảm từ 7% xuống 0% - là chỉ số lý tưởng cho việc biểu hiện protein ngoại lai. Đặc biệt, sau khi đã tối ưu, tỉ lệ GC trong phân tử thay đổi không đáng kể (tăng 1,35%) và vẫn nằm trong khoảng lý tưởng.
| Trước khi tối ưu | Sau khi tối ưu | Thông số lý tưởng | |
| CAI (*) | 0,8 | 0,92 | 0,8-1,0 |
| CFD (%) (**) | 7 | 0 | <30% |
| Tỉ lệ GC (%) | 34,39 | 35,74 | 30-70 |
Cấu trúc vector tái tổ hợp pPICzαA-kgf
Các khuẩn lạc cho kết quả PCR dương tính với vạch DNA có kích thước tương ứng với kích thước chứng dương gen kgf (468 bp) trên bản điện di được tách plasmid tái tổ hợp bằng phương pháp SDS-kiềm và kiểm tra sự gắn chèn của gen kgf lên plasmid pPICZαA bằng phản ứng PCR với cặp mồi KGF-F và AOX-R.
Kết quả điện di kiểm tra sản phẩm PCR với khuôn mẫu là plasmid thu nhận từ các khuẩn lạc dương tính ở các giếng 3, 4, 5, 6 ( Figure 2 ) cho thấy có 3 giếng xuất hiện vạch mục tiêu nằm giữa vạch 600 bp và 800 bp của thang DNA (giếng 3, 4, 6) chứng tỏ các plasmid tái tổ hợp ở các thể biến nạp này có gen kgf chèn đúng vị trí mong muốn như thiết kế. Ngược lại, ở giếng 2, sản phẩm PCR với khuôn là plasmid pPICZαA nên không có vị trí bám đặc hiệu cho mồi KGF-F, do đó sản phẩm không xuất hiện vạch. Thêm vào đó, kết quả giải trình tự cho thấy gen kgf dòng hóa tương đồng 100% về mặt trình tự so với thiết kế ban đầu và đồng khung dịch mã với trình tự α-MF. Như vậy, nghiên cứu này đã cấu trúc thành công dòng tế bào E. coli DH5α mang plasmid tái tổ hợp chứa gen mã hóa cho protein KGF.
Figure 2 . Kiểm tra plasmid tái tổ hợp bằng phương pháp PCR (mồi KGF-F và AOX-R). T: Thang DNA; 1: Chứng âm phản ứng PCR; 2: Sản phẩm PCR plasmid pPICZαA; 3 - 6. Sản phẩm PCR các plasmid tái tổ hợp.

Tạo dòng nấm men P. pastoris X33 mang gen mã hóa protein KGF
Gen AOX1 chịu trách nhiệm chính cho cơ chế sử dụng methanol ở nấm men P. pastoris , vì vậy sự sát nhập plasmid tái tổ hợp vào vùng gen này có thể xảy ra hai trường hợp: tái tổ hợp theo cơ chế chèn gen vẫn giữ nguyên vùng gen AOX1 nội sinh của nấm men, giúp chủng có khả năng sinh trưởng tốt trên môi trường có methanol (chủng mang kiểu hình Mut+) hoặc tái tổ hợp theo cơ thế thay thế, loại bỏ hoàn toàn vùng gen AOX1 nội sinh của nấm men, khiến chủng sinh trưởng yếu hơn trên môi trường có methanol (chủng mang kiểu hình MutS). Kiểu hình của chủng nấm men tái tổ hợp được kiểm tra bằng phương pháp PCR với cặp mồi AOX1 F/R. Kết quả phân tích sản phẩm PCR ( Figure 3 ) cho thấy trong số 4 thể biến nạp tiến hành kiểm tra DNA bộ gen (giếng 5, 6, 7, 8), thể biến nạp ở giếng 8 có xuất hiện 2 vạch mục tiêu theo dự đoán là vạch gen AOX1 nội sinh của nấm men với kích thước 2,2 kb và vạch gen AOX1 có chèn gen mục tiêu kgf nằm trên plasmid pPICZαA với kích thước 965 bp. Chủng nấm men tái tổ hợp này mang kiểu hình Mut + và được dự đoán có khả năng sinh trưởng tốt trên môi trường có methanol. Trường hợp chủng nấm men chỉ mang vector pPICzαA không chèn gen (Giếng 3), kết quả PCR với cặp mồi AOX sẽ cho vạch nội sinh trên bộ gen nấm men với kích thước 2,2 kb và đoạn khuếch đại nằm trong vùng gen AOX của vector với kích thước 589 bp.
Figure 3 . Kiểm tra DNA bộ gen nấm men tái tổ hợp bằng phương pháp PCR (mồi AOX-F/R). T: Thang DNA; 1. Chứng âm phản ứng PCR; 2. Sản phẩm PCR DNA bộ gen P. pastoris X33; 3. Sản phẩm PCR DNA bộ gen P. pastoris X33/pPICZαA; 4. Sản phẩm PCR plasmid tái tổ hợp pPICZαA/kgf; 5 - 9. Sản phẩm PCR DNA bộ gen của nấm men tái tổhợp P. pastoris X33::kgf

Kiểm tra khả năng biểu hiện KGF dạng tiết của chủng nấm men P. pastoris X33::kgf
Chủng nấm men tái tổ hợp P. pastoris X33:: kgf có kiểu hình Mut + được tiến hành nuôi cấy và cảm ứng biểu hiện bằng methanol nồng độ cuối 0,5%, tiến hành biểu hiện đồng thời với chủng P. pastoris X33 và P. pastoris X33/pPICZαA làm đối chứng. Kết quả kiểm tra khả năng tăng trưởng trong môi trường BMMY với methanol là nguồn carbon duy nhất ( Figure 4 A) cho thấy chủng tái tổ hợp P. pastoris X33:: kgf có khả năng sử dụng methanol để tăng sinh khối, tuy nhiên tốc độ tăng trưởng của chủng nấm men tái tổ hợp thấp hơn so với 2 chủng đối chứng là P. pastoris X33 và P. pastoris X33::pPICzαA
Figure 4 . Đánh giá biểu hiện của chủng nấm men tái tổ hợp. (A) Đường cong tăng trưởng của các chủng nấm men trong môi trường BMMY. (B) Kết quả điện di SDS-PAGE và (C) Kết quả lai western các mẫu dịch biểu hiện. T: Thang protein; 1: Chủng P. pastoris X33 có cảm ứng mathanol; 2: Chủng P. pastoris X33/pPICZαA có cảm ứng mathanol;3: Chủng P. pastoris X33::kgf không cảm ứng methanol; 4: Chủng P. pastoris X33::kgf có cảm ứng methanol.

Kết quả điện di SDS-PAGE mẫu dịch biểu hiện từ P. pastoris X33:: kgf cho thấy ngoài vạch nội sinh của chủng P. pastoris , có xuất hiện một vạch kích thước khoảng 16 kDa, dự đoán đây là vạch protein KGF mục tiêu ( Figure 4 B, giếng 4). Nhằm khẳng định protein biểu hiện chính là KGF, tiến hành lai Western với kháng thể đặc hiệu kháng KGF. Kết quả cho thấy trên màng lai xuất hiện vạch tín hiệu tương ứng với vạch proteincó kích thước 16 kDa trên bảng điện di SDS-PAGE mẫu dịch biểu hiện của chủng P. pastoris X33:: kgf được cảm ứng bởi methanol ( Figure 4 C, giếng 4). Các giếng còn lại không xuất hiện vạch tín hiệu này. Bên cạnh đó, ngoài vạch protein có kích thước 16 kDa như dự đoán thì không xuất hiện vạch kích thước nào khác. Điều này chứng tỏ protein biểu hiện dạng tiết thu được ở dịch môi trường chính là protein KGF mục tiêu và chúng tôi đã cấu trúc thành công chủng P. pastoris X33:: kgf có khả năng biểu hiện protein KGF dạng tiết.
Tinh chế và thu nhận protein KGF tái tổ hợp bằng sắc ký ái lực Heparin
50ml dịch môi trường sau biểu hiện được cho qua cột heparin 5ml với tốc độ ổn định toàn bộ quy trình là 5 ml/phút, sau đó tiến hành rửa loại tạp bằng 100%A và 30%B (Phân đoạn W - wash), cuối cùng rotein KGF được dung ly ở phân đoạn 50%B và 100% B với thể tích 15ml (Phân đoạn E – elution). Kết quả điện di SDS-PAGE và định lượng nồng độ protein bằng phương pháp Bradford cho thấy từ 50ml dịch biểu hiện, với quy trình tinh chế trên đã thu được 67 µg KGF có độ tinh sạch 99,89%, phân tích bằng phần mềm gel analyzer ( Figure 5 , giếng E 50%B), ( Table 2 ).
Figure 5 . Đánh giá kết quả tinh chế KGF tái tổ hợp bằng phương pháp SDS-PAGE. T: thang protein (GE-Healthcare); S: Mẫu dịch biểu hiện; F: Phân đoạn qua cột; W: Phân đoạn rửa cột; E: Phân đoạn dung ly

| Phân đoạn tinh chế | Thể tích (ml) | Nồng độ (μg/ml) | Độ tinh sạch (%) | KGF thu nhận (μg) | |
| Tổng | KGF | ||||
| Mẫu biểu hiện (S) | 50 | 116,71 | 18,46 | 15,82 | 791 |
| Dung ly (E) | 15 | 4,51 | 4,50 | 99,89 | 67,5 |
Hiệu suất thu hồi ở nghiên cứu này thấp hơn so với nghiên cứu tương tự trên thế giới, cụ thể là nghiên cứu của Bahadori và cộng sự năm 2018 (1,3mg < 3mg) 19 . Sự khác biệt trên có thể nằm ở quy trình biểu hiện thu nhận protein KGF từ nấm men P. pastoris. Ở nghiên cứu của Bahadori năm 2018, tác giả tiến hành thu nhận KGF ở thời điểm 96 giờ trong khi nghiên cứu này thu nhận ở 72 giờ, đồng thời lượng tế bào khởi điểm trong nghiên cứu của Bahadori cao hơn gấp 1,5 lần so với nghiên cứu này. Kết quả trên cho thấy cần có thêm nhiều nghiên cứu nhằm tối ưu điều kiện biểu hiện nhằm hướng tới việc sản xuất KGF tái tổ hợp đạt hiệu quả cao.
Thử nghiệm hoạt tính sinh học của KGF tái tổ hợp
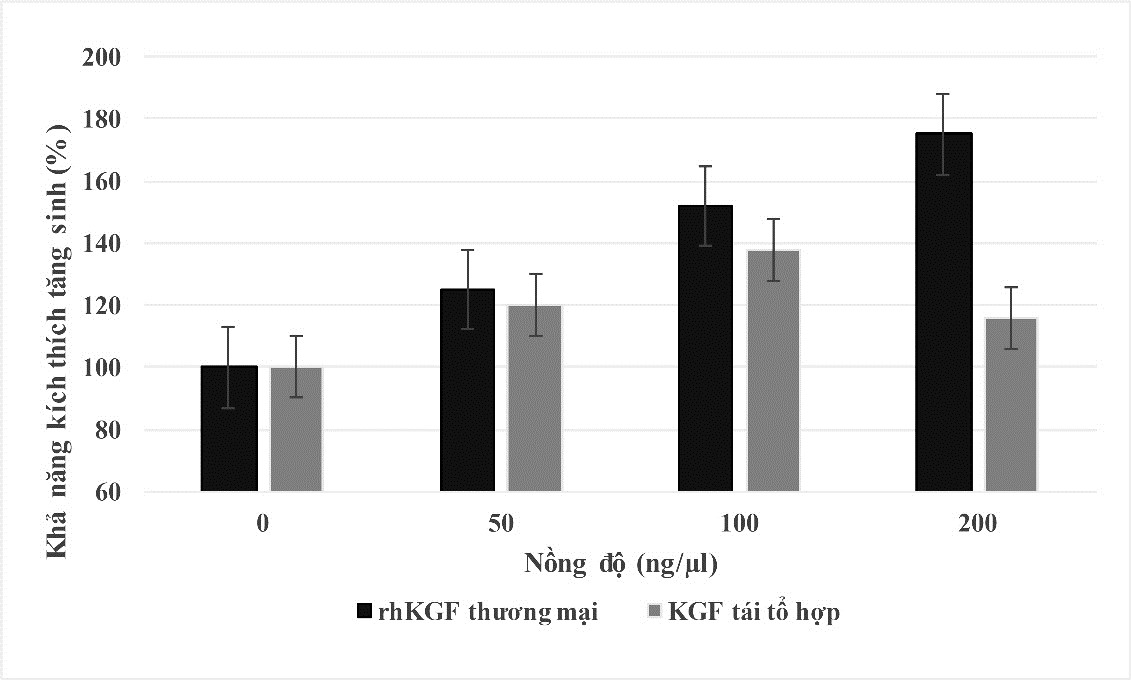
Trong tự nhiên, KGF có khả năng tương tác đặc hiệu với các tế bào biểu mô thông qua thụ thể KGF (KGF-receptor), từ đó hình thành một chuỗi phản ứng, kích hoạt các con đường dẫn truyền tín hiệu KGF và thúc đẩy quả trình tăng sinh, biệt hóa tế bào. Nghiên trước đây cho thấy KGF kích thích tăng sinh tế bào A549 thông qua nhận diện thụ thể KGRF trên bề mặt tế bào này 19 . Ở nghiên cứu này, trong điều kiện không có tế bào keratinocyte chúng tôi kiểm tra cấu hình tự nhiên của protein KGF tái tổ hợp thông qua việc nhận diện thụ thể KGFR trên tế bào A549. Kết quả của thử nghiệm ( Figure 6 ) cho thấy mẫu KGF thu nhận từ dịch biểu hiện P. pastoris có hoạt tính kích thích tăng sinh 1,2 lần ở nồng độ 50 ng/mL, 1,38 lần ở nồng độ 100 ng/mL, trong khi mẫu chứng dương là KGF thương mại biểu hiện từ E. coli (R & D systems) có hoạt tính kích thích tăng sinh ở nồng độ 50 ng/ml là 1,25 lần và tăng dần đến 1,52 lần ở nồng độ 100 ng/ml. Tuy nhiên, ở nồng độ 200 ng/mL, KGF thương mại cho hoạt tính kích thích tăng sinh 1,75 lần, trong khi KGF tái tổ hợp chỉ cho khả năng kích thích tăng sinh 1,16 lần, thấp hơn so với nồng độ 50 ng/ml và 100 ng/ml. Kết quả này có thể do sự khác biệt về dung dịch trữ giữa KGF thương mại và KGF tái tổ hợp, khác biệt về thành phần muối trong dung môi có thể gây ra sự kết cụm của các phân tử KGF tái tổ hợp khi sử dụng ở nồng độ cao và từ đó làm giảm tác động của KGF tái tổ hợp 20 . Kết quả thực nghiệm này cho thấy cần tiến hành thêm các nghiên cứu nhằm khảo sát nồng độ sử dụng tối ưu của KGF tái tổ hợp trong kích thích tăng sinh tế bào, cũng như khảo sát dung dịch lưu trữ KGF tái tổ hợp.
Figure 6 . Hoạt tính sinh học của KGF tái tổ hợp trên dòng tế bào A549. Chứng âm (PBS) được xem có mức độ tăng sinh 100%. *: p-value<0,05; **:p-value<0,01
KẾT LUẬN
Kết quả nghiên cứu trên cho thấy chúng tôi đã tạo thành công dòng nấm men P. pastoris tái tổ hợp mang gen mã hóa cho protein KGF. Dưới sự kiểm soát của promoter AOX1, protein KGF tái tổ hợp được biểu hiện dưới dạng tiết và được tinh sạch bằng phương pháp sắc ký ái lực Heparin với độ tinh sạch cao (> 99%) cũng như hoạt tính kích thích tăng sinh tế bào tương tự như KGF tự nhiên. Với mục tiêu ứng dụng KGF trong lĩnh vực dược phẩm và mỹ phẩm ở quy mô lớn và giảm giá thành sản phẩm, cần tăng quy mô biểu hiện và tối ưu hóa các điều kiện nuôi cấy có kiểm soát, cũng như xây dựng quy trình thử nghiệm hoạt tính sinh học trên nhiều mô hình khác nhau.
DANH MỤC TỪ VIẾT TẮT
AOX : Alcohol oxidase
FGF : Fibroblast Growth Factor
KGF : Keratinocyte Growth Factor/ Fibroblast Growth Factor – 7
KGFR : Keratinocyte Growth Factor Receptor
LB : Luria-Bertani
MTT : Tetrazolium (3- [4,5- dimethylthiazol- 2- yl-] 2,5- diphenyltetrazolium bromide
PCR : Polymerase Chain Reaction
SDS-PAGE : Sodium dodecyl sulfate – polyacrylamide gel electrophoresis
XUNG ĐỘT LỢI ÍCH
Các tác giả cam đoan rằng họ không có xung đột lợi ích.
ĐÓNG GÓP CỦA TÁC GIẢ
Nghiên cứu này được thiết kế bởi tác giả Đặng Thị Phương Thảo và Nguyễn Hiếu Nghĩa. Tác giả Nguyễn Phạm Anh Thư và Nguyễn Thị Thùy Trang tiến hành thu thập số liệu tại phòng thí nghiệm và xử lý kết quả. Tác giả Đăng Thị Phương Thảo và tác giả Nguyễn Phạm Anh Thư tham gia viết bản thảo.
LỜI CÁM ƠN
Nghiên cứu được tài trợ bởi Đại học Khoa học Tự nhiên, Đại học Quốc gia Thành phố Hồ Chí Minh (ĐHQG-HCM) trong khuôn khổ đề tài mã số T2019-22
References
- Rubin J.S., Osada H., Finch P.W., Taylor W.G., Rudikoff S., Aaronson S.A.. Purification and characterization of a newly identified growth factor specific for epithelial cells. Proceedings of the National Academy of Sciences. 1989;86(3):802-806. PubMed Google Scholar
- Finch P.W., Rubin J.S.. Keratinocyte growth factor/fibroblast growth factor 7, a homeostatic factor with therapeutic potential for epithelial protection and repair. Advances in cancer research. 2004;91:70-137. Google Scholar
- Rubin J.S., Bottaro D.P.. Keratinocyte growth factor. Cell biology international. 1995;19(5):399-412. PubMed Google Scholar
- Marchese C., Messina A., Faggioni A.. Human keratinocyte growth factor activity on proliferation and differentiation of human keratinocytes: differentiation response distinguishes KGF from EGF family. Journal of cellular physiology. 1990;144(2):326-332. PubMed Google Scholar
- Peng Y., Wu S., Tang Q., Li S., Peng C.. KGF-1 accelerates wound contraction through the TGF-β1/Smad signaling pathway in a double-paracrine manner. Journal of Biological Chemistry. 2019;294(21):8361-8370. PubMed Google Scholar
- Feng Z.G., Pang S.F.. Recombinant keratinocyte growth factor 1 in tobacco potentially promotes wound healing in diabetic rats. BioMed research international. 2014;;:. PubMed Google Scholar
- Araújo R., Lôbo M., Trindade K., Silva D.F., Pereira N.. Fibroblast Growth Factors: A Controlling Mechanism of Skin Aging. Skin pharmacology and physiology. 2019;4(5):275-282. PubMed Google Scholar
- Ceccarelli S., Romano F., Angeloni A., Marchese C.. Potential dual role of KGF/KGFR as a target option in novel therapeutic strategies for the treatment of cancers and mucosal damages. Expert opinion on therapeutic targets. 2012;16(4):377-393. PubMed Google Scholar
- Finch P.W., Rubin J.S.. Keratinocyte growth factor expression and activity in cancer: implications for use in patients with solid tumors. Journal of the National Cancer Institute. 2006;98(12):812-824. PubMed Google Scholar
- Gegechkori N., Haines L., J.J. Lin. Long Term and Latent Side Effects of Specific Cancer Types. The Medical clinics of North America. 2017;101(6):1053. PubMed Google Scholar
- Barasch A., Epstein J., Tilashalski K.. Palifermin for management of treatment-induced oral mucositis in cancer patients. Biologics: targets & therapy. 2009;3:111. PubMed Google Scholar
- Ron D., Bottaro D.P., Finch P.W., Morris D., Rubin J.S., Aaronson S.. Expression of biologically active recombinant keratinocyte growth factor. Structure/function analysis of amino-terminal truncation mutants. Journal of Biological Chemistry. 1993;268(4):2984-2988. Google Scholar
- Luo Y., Cho H.H., Jones R.B., Jin C., McKeehan W.L.. Improved production of recombinant fibroblast growth factor 7 (FGF7/KGF) from bacteria in high magnesium chloride. Protein expression and purification. 2004;33(2):326-331. PubMed Google Scholar
- Han S.Y., Jin C.Y.. Production of Recombinant Human Keratinocyte Growth Factor from Bombyx mori (Lepidopera: Bombycidae) Bm5 Cells. Journal of Life Science. 2011;21(6):907-911. Google Scholar
- Hsu Y.R., Hsu E.W.. Human keratinocyte growth factor recombinantly expressed in Chinese hamster ovary cells: isolation of isoforms and characterization of post-translational modifications. . Protein expression and purification. 1998;12(2):189-200. PubMed Google Scholar
- Ahmad M., Hirz M., Pichler H., Schwab H.. Protein expression in Pichia pastoris: recent achievements and perspectives for heterologous protein production. Applied microbiology and biotechnology. 2014;98(12):5301-5317. PubMed Google Scholar
- El-Mansi E., Nielsen J., Mousdale D., Carlson R.P.. Metabolic analysis and optimization of microbail and animal cell bioprocesses, Fermentation microbiology and biotechnology. Second ed: CRC press. 2018;:. PubMed Google Scholar
- Osslund T.D., Syed R.. Correlation between the 1.6 Å crystal structure and mutational analysis of keratinocyte growth factor. Protein Science. 1998;7(8):1681-1690. PubMed Google Scholar
- Bahadori Z., Kalhor H.R., S.J. Mowla. Producing functional recombinant human keratinocyte growth factor in Pichia pastoris and investigating its protective role against irradiation. Enzyme and microbial technology. 2018;111:12-20. PubMed Google Scholar
- WeiWang Chrostopher J.R.. Aggregration of therapeutic proteins. Wiley, A Jhn Wiley & Sons, Inc, Publication. 2010;:369. Google Scholar





