
 Open Access
Open Access Abstract
Antibiotic resistance has emerged and persisted as one of the major threats to public health and the sustainable development of healthcare systems worldwide in recent years. The primary reason why antibiotic resistance is regarded as a global medical crisis lies in the development of new antibiotics is not keeping pace with the emergence of antibiotic-resistant strains. Currently, one of the proposed solutions to this crisis, as suggested by scientists, is the investigation of alternative antimicrobial agents that exhibit high efficacy while also possessing characteristics that limit the emergence of resistance in target pathogenic strains. Among such alternatives, endolysins, a class of enzymes derived from bacteriophages, have garnered considerable research interest. These enzymes are utilized by bacteriophages to degrade the peptidoglycan layer of bacterial cell walls during the infection process. The ability of endolysins to lyse bacterial cells when applied externally, combined with their high specificity at the genus or species level and a low propensity for inducing resistance, renders them promising candidates as substitutes for conventional antibiotics. This review aims to outline the structural features and mechanisms of action of endolysins, and to discuss in depth their characteristics and potential applications across various fields.
Giới thiệu
Tình trạng kháng kháng sinh đã được Tổ chức Y tế Thế giới WHO nhận định là một trong những mối đe dọa lớn nhất đối với sức khỏe con người, an ninh lương thực và sự phát triển của thế giới 1 . Số lượng các ca bệnh nhiễm vi sinh vật kháng kháng sinh, đặc biệt là các chủng kháng đa thuốc liên tục được ghi nhận ở mức cao ở cả Việt Nam và các nước trên thế giới 2 , 3 , 4 , 5 . Các chủng kháng kháng sinh gây nhiều khó khăn trong công tác điều trị, khiến sức khỏe người nhiễm bị ảnh hưởng nghiêm trọng và khả năng tử vong tăng cao 2 .
Việc kiểm soát tình trạng kháng kháng sinh hiện nay chủ yếu tập trung vào việc hạn chế sử dụng kháng sinh truyền thống. Các chiến lược có thể được đề cập đến như tiêm vaccine nhằm phòng ngừa sự lây nhiễm hay sử dụng các biện pháp kháng khuẩn từ probiotic hay các chất kháng khuẩn thay thế. Vaccine ngừa vi khuẩn gây bệnh vẫn luôn là biện pháp ngăn cản sự lây lan mầm bệnh hiệu quả, từ đó ngăn chặn sự phổ biến của các chủng mang gen kháng kháng sinh. Tuy nhiên, thách thức lớn nhất trong việc sử dụng vaccine để kiểm soát tình trạng kháng kháng sinh nằm ở việc lựa chọn được kháng nguyên vaccine có phổ rộng, tính bảo tồn cao và có khả năng tạo đáp ứng miễn dịch kéo dài 6 . Cùng mang giá trị trong giảm thiểu sự lây nhiễm của vi khuẩn gây bệnh là việc sử dụng các vi sinh vật sống có lợi – probiotic. Probiotic thường được ứng dụng trong bảo quản các sản phẩm nông nghiệp và thực phẩm, cũng như trong việc nâng cao sức khỏe của người và vật nuôi, nhờ khả năng ngăn chặn sự lây nhiễm của vi sinh gây bệnh bằng tương tác cạnh tranh hay kích thích phản ứng miễn dịch của vật chủ. Các chủng probiotic mới liên tục được tìm ra, gần đây nhất là chủng Bacillus mới trong khám phá của Golnari và cộng sự với khả năng kháng phổ rộng nhiều chủng vi khuẩn gây bệnh 7 . Trên thực tế, các phương pháp kháng khuẩn bằng probiotic có tốc độ phát triển tương đối chậm bởi lo ngại trong việc truyền ngang gen kháng kháng sinh từ chủng probiotic sang chủng vi khuẩn gây bệnh khi chúng sinh trưởng trong cùng môi trường, từ đó yêu cầu phương pháp sàng lọc chủng mới tiên tiến 8 . Khác với hai chiến lược trên, sử dụng các chất kháng khuẩn thay thế giúp trực tiếp tiêu diệt vi khuẩn gây hại mục tiêu. Hai trong số các biện pháp đang được nghiên cứu phổ biến là sử dụng thể thực khuẩn và các peptide kháng khuẩn. Các thể thực khuẩn trong những năm gần đây đang được đẩy mạnh nghiên cứu để trở thành thành phần trong các thuốc kháng khuẩn thế hệ mới. Tuy nhiên, trái với kỳ vọng, các thử nghiệm lâm sàng về thuốc chứa thực khuẩn thể đều không mang lại kết quả thuận lợi 6 . Các peptide kháng khuẩn hiện nay vẫn đang là đối tượng tiềm năng với nhiều nghiên cứu lâm sàng có kết quả thuận lợi trong công cuộc tìm kiếm chất kháng khuẩn thay thế của ngành khoa học y khoa 1 .
Endolysin, một loại enzyme từ thực khuẩn thể với khả năng ly giải tế bào vi khuẩn, là một chất kháng khuẩn thay thế đang dần được đẩy mạnh nghiên cứu trong hai thập kỷ trở lại đây 9 . Endolysin là các enzyme thủy phân peptidoglycan (peptidoglycan hydrolase) được thực khuẩn thể tổng hợp và sử dụng vào cuối chu trình sinh tan để phá cấu trúc thành tế bào vi khuẩn chủ, khiến tế bào vỡ do sốc thẩm thấu, từ đó giúp giải phóng thế hệ thực khuẩn thể tiếp theo. Mặc dù được thực khuẩn thể sử dụng để ly giải lớp peptidoglycan từ bên trong tế bào chủ, endolysin đã được chứng minh vẫn mang khả năng ly giải khi tác động từ bên ngoài. Điều này khiến enzyme này được xem như một chất kháng khuẩn tiềm năng, và nhiều nghiên cứu trên thế giới cũng đã ứng dụng thành công endolysin dù ở dạng tinh sạch hay tái tổ hợp 10 , 11 . Mặt khác, lớp peptidoglycan là thành phần chính cấu trúc nên thành tế bào vi khuẩn, có tính bảo tồn cao và ít bị biến đổi; vì vậy, rất ít trường hợp kháng endolysin được ghi nhận và các trường hợp mang khả năng kháng thường bị suy giảm độc lực 11 . Một đặc điểm nổi trội khác của endolysin là chúng mang khả năng ly giải đặc hiệu đối với các thành viên trong cùng chi, loài hoặc kiểu huyết thanh. Đặc điểm này giúp hạn chế sự ảnh hưởng đến hệ vi sinh vật trong môi trường tác động, giảm thiểu khả năng hình thành tính kháng trên các loài vi khuẩn sống cùng môi trường với loài vi khuẩn mục tiêu 10 , 12 , 13 .
Cùng với những nỗ lực đẩy lùi tình trạng kháng kháng sinh trên toàn thế giới, việc sở hữu các đặc điểm thuận lợi để trở thành một chất kháng khuẩn thay thế cho kháng sinh truyền thống giúp số lượng các nghiên cứu về ứng dụng endolysin trong an toàn thực phẩm, xử lý môi trường, y tế,… không ngừng tăng lên. Nhằm cung cấp các thông tin cần thiết giúp đẩy mạnh các nghiên cứu về endolysin, bài tổng quan này tập trung giới thiệu các đặc điểm về cấu trúc và cơ chế hoạt động của enzyme kháng khuẩn này, cùng với đó là các đặc điểm của endolysin khi được sử dụng như một chất kháng khuẩn và thông tin về ứng dụng của endolysin trong một số lĩnh vực.
Cấu trúc VÀ CƠ CHẾ HOẠT ĐỘNG của endolysin
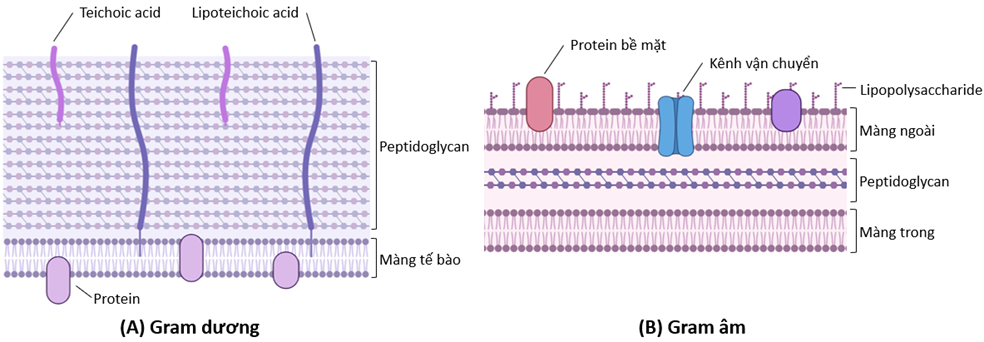
Thành tế bào vi khuẩn – mục tiêu ly giải của endolysin – mang những đặc điểm khác nhau giữa vi khuẩn Gram dương và vi khuẩn Gram âm ( Figure 1 ). Tế bào vi khuẩn Gram dương được bảo vệ bởi lớp peptidoglycan dày (khoảng 20-80 nm), cùng các phân tử carbonhydrate và các protein được đính cộng hóa trị trên bề mặt 10 . Bên dưới lớp peptidoglycan là lớp màng đôi phospholipid bao bọc tế bào chất. Mặt khác, thành tế bào vi khuẩn Gram âm bao gồm lớp peptidoglycan mỏng hơn (khoảng 5-10 nm) nằm giữa hai lớp màng đôi: màng trong bao bọc tế bào chất; màng ngoài mang lớp lipopolysaccharide, các kênh vận chuyển, và các thụ thể đính màng, đóng vai trò là lớp bảo vệ đầu tiên của tế bào khỏi các tác nhân bên ngoài. Bởi những sự khác biệt trên nên các đặc điểm cấu trúc của endolysin cũng phụ thuộc vào mục tiêu ly giải của chúng: thành tế bào vi khuẩn Gram dương hay vi khuẩn Gram âm.
Figure 1 . Cấu trúc thành tế bào vi khuẩn Gram dương (A) và vi khuẩn Gram âm (B) (tạo bằng phần mềm biorender.com)
Cấu trúc endolysin vi khuẩn Gram dương
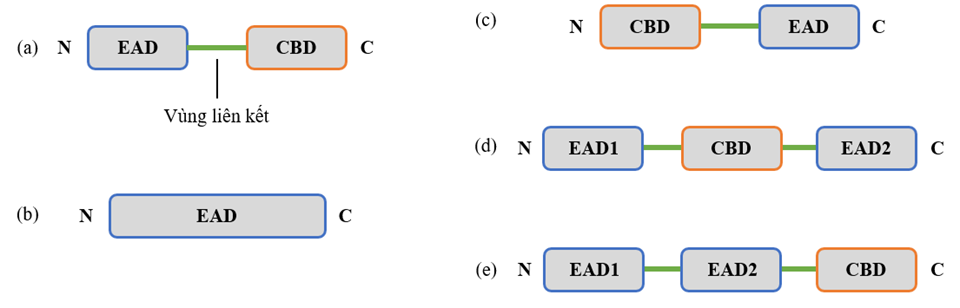
Các endolysin có mục tiêu là thành tế bào vi khuẩn Gram dương thường có cấu trúc dạng mô đun bao gồm từ hai miền (domain) trở lên, các miền này được chia thành hai nhóm chức năng chính: miền mang chức năng enzyme (enzymatically active domain – EAD) nằm về phía đầu N của protein và miền bám màng (cell-wall binding domain – CBD) nằm về phía đầu C 14 , 15 ( Figure 2 a). Các EAD mang nhiệm vụ phân cắt các liên kết trong cấu trúc lớp peptidoglycan. Trong khi đó, các CBD mang vai trò nhận diện và gắn với các thành phần trên thành tế bào, giúp định hướng endolysin đến lớp peptidoglycan. Endolysin vi khuẩn Gram dương thường bao gồm một CBD và tối thiểu một EAD, các EAD và CBD được liên kết với nhau bởi các vùng liên kết (linker region) linh động 16 .
Cấu trúc endolysin vi khuẩn Gram âm
Khác với các endolysin vi khuẩn Gram dương, endolysin từ thực khuẩn thể vi khuẩn Gram âm thường có cấu trúc hình cầu và thường chỉ gồm một EAD 17 , 18 ( Figure 2 b). Sự vắng mặt của CBD có thể được lý giải là do lớp màng ngoài đóng vai trò ngăn các phân tử endolysin từ các tế bào đã bị ly giải tiếp xúc với lớp peptidoglycan trên tế bào chưa bị xâm nhiễm, vì vậy trong trường hợp này, sự có mặt của CBD được xem như vô nghĩa 15 . Tuy nhiên, một số trường hợp endolysin vi khuẩn Gram âm có cấu trúc mô đun gồm từ hai miền trở lên đã được các nhà khoa học ghi nhận. Các endolysin mang cấu trúc mô đun thường có sự đa dạng trong thành phần cũng như cách sắp xếp các miền. Ngoài ra, một điểm khác biệt nữa so với endolysin vi khuẩn Gram dương là ở endolysin vi khuẩn Gram âm, các EAD thường nằm về phía đầu C của protein và các CBD nằm về phía đầu N. Một số ví dụ điển hình cho các endolysin nói trên là hai endolysin KZ144 và EL188 từ thực khuẩn thể chi Pseudomonas mang cấu trúc gồm một EAD nằm ở đầu C và một CBD nằm về đầu N 18 ( Figure 2 c); endolysin từ thực khuẩn thể λSA2 nhóm streptococcus mang một CBD xen giữa hai EAD 19 ( Figure 2 d); và cấu trúc gồm một CBD nằm về đầu C và hai EAD nằm về đầu N thường thấy ở endolysin từ thực khuẩn thể nhóm staphylococcus 15 ( Figure 2 e).
Figure 2 . Cấu trúc một số endolysin điển hình. (a) Cấu trúc gồm một miền chức năng enzyme (EAD) đầu N và một miền bám màng (CBD) đầu C; (b) Cấu trúc chỉ gồm EAD; (c) Cấu trúc gồm một CBD đầu N và một EAD đầu C; (d) Cấu trúc gồm một CBD xen giữa hai EAD; (e) Cấu trúc gồm một CBD đầu C và hai EAD đầu N.
Miền mang chức năng enzyme – EAD
Các endolysin được phân loại dựa vào hoạt tính enzyme của EAD. Dựa vào loại liên kết mục tiêu, EAD được phân thành bốn lớp chính: glycosidase, amidase, endopeptidase và lytic transglycosylase.
Các glycosidase mang khả năng phân cắt liên kết β-1,4 glycosidic giữa N-acetylmuramic acid (MurNAc) và N-acetylglucosamine (GlcNAc) trong mạch polymer chính của lớp peptidoglycan. Một trong hai lớp phụ của glycosidase, N-acetylglucosaminidase, phân cắt thành phần glycan thuộc phân tử GlcNAc. Một ví dụ điển hình cho hoạt tính này là endolysin từ thực khuẩn thể λSA2 nhóm streptococcus 19 . Ngược lại, lớp phụ thứ hai của glycosidase là N-acetylmuramidase phân cắt thành phần glycan thuộc phân tử MurNAc. Endolysin Cpl-1 nhóm pneumococcus 20 và B30 nhóm streptococcus 21 là các glycosidase mang hoạt tính này.
Lớp EAD thứ hai gồm N-acetylmuramoyl-L-alanine amidase có bản chất là một amidohydrolase (enzyme thủy phân liên kết amine). Lớp enzyme này mang khả năng phân cắt liên kết giữa phân tử MurNAc trong mạch chính peptidoglycan và L-alanine, amino acid đầu tiên trong mạch nhánh peptide. Một số ví dụ điển hình cho lớp này bao gồm endolysin LysK từ thực khuẩn thể K nhóm staphylococcus 22 và endolysin Ply511 12 , endolysin PlyPSA 16 từ thực khuẩn thể chi Listeria .
Mục tiêu của lớp EAD thứ ba, endopeptidase, là liên kết giữa hai amino acid trên mạch nhánh peptide trong cấu trúc peptidoglycan. Loại endolysin này có thể phân cắt nội liên kết thuộc mạch nhánh peptide hoặc liên kết giữa hai mạch nhánh peptide liền kề. Một số ví dụ điển hình cho loại này là Ply500 L-alanyl-D-glutamate endolysin với mục tiêu ly giải là chi Listeria 12 hay Ф11 D-alanyl-glycyl endolysin ly giải nhóm staphylococcus 23 .
Lớp EAD cuối cùng là lytic transglycosylase. Khác với ba nhóm còn lại, lytic transglycosylase không mang hoạt tính thủy phân vì chúng không cần phân tử nước để thực hiện hoạt động enzyme của mình 24 . Hoạt tính của lytic transglycosylase tương tự các glycosidase với khả năng phân cắt liên kết β-1,4 glycosidic giữa MurNAc và GlcNAc trên mạch peptidoglycan chính. Endolysin gp144 từ thực khuẩn thể ФKZ là một trong những endolysin được xác định mang hoạt tính lytic transglycosylase 25 .
Miền bám màng – CBD
Miền bám màng CBD mang chức năng gắn và giữ endolysin với thành tế bào thông qua các liên kết không cộng hóa trị với một số thành phần trên thành, chẳng hạn như phối tử murein (murein ligand) hay các chuỗi polymer thứ cấp trong cấu trúc lớp peptidoglycan như teichoic acid hay polychaccharide 15 . Bên cạnh chức năng liên kết với mục tiêu ly giải, các CBD còn chịu trách nhiệm chính cho tính đặc hiệu của endolysin. Ở hầu hết trường hợp, phổ nhận diện của CBD bao gồm thành viên trong cùng một chi 15 . Tuy nhiên, một số ngoại lệ đã được ghi nhận với CBD có phổ nhận diện hẹp hay rộng hơn thông thường. Chẳng hạn, một số endolysin kháng chi Listeria có tính đặc hiệu lên đến mức kiểu huyết thanh (serovar) hay chủng (strain) 10 ; ngược lại, endolysin từ phage ФAB2 xâm nhiễm loài Acinetobacter baumannii ( A. baumannii ) đã được ghi nhận có khả năng ly giải cả tế bào vi khuẩn Gram dương lẫn Gram âm 26 . Cuối cùng, một vai trò quan trọng khác của CBD được thể hiện ở giai đoạn sau ly giải là cố định phân tử endolysin trên các mảnh vỡ của thành tế bào vi khuẩn, giúp ngăn các endolysin tiếp xúc các tế bào lân cận chưa được xâm nhiễm bởi thực khuẩn thể 27 .
Quá trình ly giải tế bào vi khuẩn
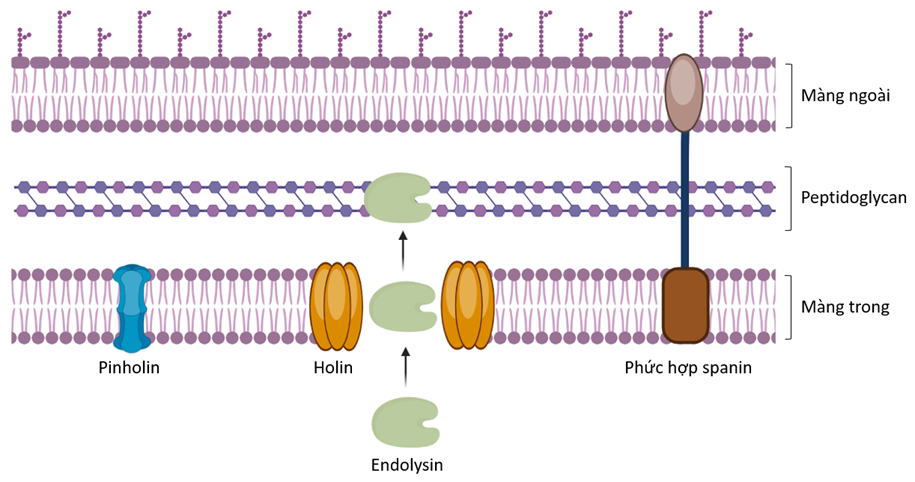
Như đã đề cập từ trước, endolysin được các thực khuẩn thể sử dụng để ly giải thành tế bào vi khuẩn chủ, từ đó giúp giải phóng thế hệ thực khuẩn thể mới. Chi tiết hơn, quá trình ly giải tế bào chủ của các thực khuẩn thể DNA mạch đôi có thể bao gồm ba cơ chế khác nhau với sự có mặt của các phân tử chức năng khác bên cạnh endolysin được mô tả ở Figure 3 28 , 29 . Cơ chế đầu tiên bao gồm sự có mặt của holin, một protein kỵ nước có kích thước nhỏ được thực khuẩn thể tạo ra vào cuối giai đoạn xâm nhiễm. Khi được tạo ra đến nồng độ đạt ngưỡng giới hạn, các phân tử holin tiến hành quá trình tự oligomer hóa và xen qua lớp màng đôi, mở ra các lỗ lớn trên màng, từ đó cho phép endolysin đi qua và tiếp cận lớp peptidoglycan 30 . Một cơ chế khác cũng diễn ra song song, thông qua hoạt động của một phân tử cũng mang khả năng xuyên màng là pinholin. Khi được kích hoạt, các pinholin, lúc này đang tập trung tại vùng chu chất, tiến hành tự oligomer hóa tạo thành các kênh xuyên màng. Sự hình thành của các kênh này khiến điện thế màng giảm, tạo điều kiện cho các phân tử endolysin gấp cuộn thành dạng mang hoạt tính và tiến hành ly giải lớp peptidoglycan. Sau cùng, để ly giải lớp màng ngoài tế bào ở các vi khuẩn Gram âm, các thực khuẩn thể cần đến hoạt động của một protein ly giải khác được gọi là phức hợp spanin. Phức hợp spanin hai thành phần bao gồm một lipoprotein có kích thước nhỏ thuộc màng ngoài vi khuẩn (o-spanin) và một protein xuyên màng tế bào chất (i-spanin). Chức năng phá vỡ màng ngoài của spanin chủ yếu được thực hiện thông qua cơ chế dung hợp màng trong màng ngoài, từ đó mở ra các lỗ lớn giải phóng thế hệ thực khuẩn thể mới ra môi trường ngoại bào 31 .
Figure 3 . Endolysin và các protein hỗ trợ tham gia vào quá trình ly giải tế bào vi khuẩn Gram âm (tạo bằng công cụ biorender.com)
endolysin TRONG VAI TRÒ LÀ MỘT CHẤT KHÁNG KHUẨN
Nghiên cứu đầu tiên về khả năng kháng khuẩn của endolysin được thực hiện bởi Frederick W. Twort vào năm 1925 khi tác giả này đưa ra giả thuyết về vai trò quan trọng của endolysin trong khả năng ly giải tế bào vi khuẩn của thể thực khuẩn. Endolysin, lúc này được gọi là chất ly giải vi khuẩn (bacterial lysin) tiết ra bởi thực khuẩn thể cho thấy khả năng ly giải tế bào vi khuẩn ở cả hai trạng thái sống và chết, khác với thể thực khuẩn chỉ có thể ly giải tế bào sống 32 . Giả thuyết này sau đó đã được xác nhận bởi Reynals (1926), Evans (1935) và Maxted (1957) bằng nhiều thí nghiệm khác nhau 33 , 34 , 35 . Ở bước phát triển tiếp theo, các nhà khoa học đã tinh sạch được endolysin từ thực khuẩn thể và sử dụng trong các thí nghiệm xác định đặc tính như trong nghiên cứu của Krause vào năm 1957 và sau đó là Fischetti và cộng sự vào năm 1971 36 . Từ đầu những năm 2000, endolysin đã bắt đầu được đánh giá khả năng kháng khuẩn trên các mô hình in vivo , mở đầu là nghiên cứu của Nelson và cộng sự trong thử nghiệm sử dụng endolysin từ thực khuẩn thể C1 để kháng lại sự phát triển của nhóm streptococcus trên mô hình chuột 37 . Nối tiếp là công bố của Loeffler và cộng sự về việc sử dụng endolysin Cpl-1 kháng vi khuẩn nhóm pneumococcal thông qua đường tiêm tĩnh mạch 37 . Năm 2013 đánh dấu bước phát triển tiếp theo của việc ứng dụng endolysin trong kháng khuẩn với thử nghiệm lâm sàng đầu tiên được tiến hành. Thuốc dựa trên endolysin SAL-1, được thiết kế để điều trị nhiễm trùng tụ cầu kháng kháng sinh 38 . Đến năm 2020, Staphefekt™, sản phẩm chứa endolysin đầu tiên trên thế giới dành cho việc điều trị bệnh trên người, đã được ra mắt.
Bên cạnh các nghiên cứu trên, nhiều nghiên cứu sử dụng thành công endolysin cũng đã được công bố 39 , 40 , 41 , 42 , 43 . Xa hơn, số lượng các sản phẩm có thành phần là endolysin, hoặc được thiết kế dựa trên endolysin, đang bước vào giai đoạn thử nghiệm lâm sàng hoặc đưa ra thị trường ngày một tăng lên 44 . Những minh chứng trên cho thấy endolysin đang từng bước trở thành một trong những chất kháng khuẩn thay thế cho kháng sinh truyền thống. Phần này sẽ thảo luận về các đặc tính của endolysin giúp loại enzyme này được quan tâm nghiên cứu trên thế giới như hiện nay.
Tính sinh miễn dịch của endolysin
Cơ thể có khả năng tạo ra đáp ứng miễn dịch đối với các tác nhân có bản chất là protein. Ở một số trường hợp, đáp ứng miễn dịch có thể dẫn đến những hệ quả xấu như gây ra những phản ứng quá mẫn hoặc các bệnh tự miễn, thậm chí gây tử vong. Trên thực tế, đây là một trong những nhân tố chính hạn chế ứng dụng endolysin, phân tử sinh học có bản chất là protein, như một loại thuốc điều trị nhiễm khuẩn trên người và động vật. Để giải quyết vấn đề trên, nhiều thí nghiệm kiểm tra trên mô hình động vật đã được các nhà nghiên cứu thực hiện. Cụ thể, kết quả nghiên cứu trên chuột của Witzenrath và cộng sự 45 về tính sinh miễn dịch của endolysin Cpl-1 (kháng nhóm pneumococcus) cho thấy: nguy cơ sinh các phản ứng dị ứng hoặc quá mẫn là thấp, với lượng IgE không có sự thay đổi mặc dù lượng IgG tăng cao. Các kết quả kiểm tra khác cũng cho thấy không có tác dụng phụ nào khác xảy ra, cũng như các cytokine gây viêm được giữ ở mức bình thường và phản ứng kích hoạt bổ thể không diễn ra.
Mặt khác, các phản ứng miễn dịch còn là nguyên nhân cho những nghi ngờ về hiệu quả kháng khuẩn của endolysin khi sử dụng trên cơ thể. Khi gây đáp ứng, các kháng thể được hệ miễn dịch tạo ra sẽ trung hòa các endolysin được đưa vào cơ thể, điều này có thể khiến các endolysin mất hoạt tính. Tuy nhiên, nghiên cứu sử dụng endolysin LysGH15 kháng nhóm staphylococcus trên mô hình chuột của Zhang và cộng sự đã cho kết quả đáng mong đợi 46 . Cụ thể, các kháng thể đặc hiệu được sản sinh không làm mất khả năng ly giải và khả năng bám mục tiêu của LysGH15 trong các thí nghiệm in vitro . Bên cạnh đó, với hiệu giá kháng thể ở mức cao nhất, LysGH15 vẫn giữ được khả năng bảo vệ cơ thể chuột khi cho nhiễm chủng Staphylococcus aureus kháng methicillin (MRSA). Một số nghiên cứu khác đã được thực hiện trên các endolysin PlyG kháng Bacillus anthracis và PlyC kháng Streptococcus pyogenes đều cho thấy rằng với sự có mặt của kháng thể trung hòa đặc hiệu, tốc độ tiêu diệt vi khuẩn mục tiêu của endolysin tuy giảm nhưng không dừng lại 47 .
Tính an toàn khi sử dụng endolysin như chất kháng khuẩn
Bên cạnh tính an toàn liên quan đến các phản ứng miễn dịch sản sinh khi sử dụng, các tác dụng phụ khác cũng được các nhà nghiên cứu quan tâm khi tiến hành đưa các sản phẩm endolysin vào giai đoạn thử nghiệm lâm sàng. SAL200, sản phẩm đầu tiên có thành phần là endolysin được phát triển để điều trị nhiễm khuẩn MRSA, đã được thử nghiệm tính an toàn trên nhiều đối tượng. Khi thử nghiệm tiêm tĩnh mạch trên chó và khỉ, không có tác dụng phụ nào xảy ra hoặc được báo cáo trên cả hai đối tượng thử nghiệm này 48 , 49 . Ở giai đoạn tiếp theo, tính an toàn của SAL200 cũng được đánh giá trên người. Các đặc điểm về dược động học, dược lực học và khả năng dung nạp được đánh giá trên các tình nguyện viên nam tiếp nhận SAL200 qua tiêm tĩnh mạch. Kết quả cho thấy tất cả các tình nguyện viên không gặp tác dụng phụ nào đáng kể và không bị tái nhiễm, ngoài trừ ba tình nguyện viên báo cáo về các triệu chứng đau đầu, mệt mỏi và đau cơ. Ngoài ra, ở các kết quả kiểm tra sinh hóa, huyết học và đông máu, dấu hiệu sinh tồn, khám tổng quát, không có sự bất thường nào có ý nghĩa về mặt lâm sàng được ghi nhận ở tất cả các tình nguyên viên 48 .
Khả năng hình thành tính kháng endolysin của các loài vi khuẩn
Tính đặc hiệu giới hạn trên các thành viên cùng chi, loài hoặc thậm chí kiểu huyết thanh là nhân tố chính giúp giảm thiểu sự hình thành tính kháng ở các loài vi khuẩn mục tiêu đối với endolysin. Tuy vậy, khả năng hình thành tính kháng đối với endolysin không phải là số không tuyệt đối, nhưng được đánh giá là thấp nếu so sánh với việc sử dụng các kháng sinh truyền thống 11 , 15 , 47 . Cho đến hiện tại, một số ít trường hợp mang khả năng kháng endolysin hoặc các protein lai có thành phần endolysin đã được ghi nhận, nhưng đa số các trường hợp đều đi kèm với sự giảm độc lực hoặc tăng tính nhạy cảm với các loại kháng sinh khác 11 . Trong nghiên cứu của Becker và cộng sự, quá trình phơi nhiễm liên tục với endolysin LysK ở nồng độ dưới ngưỡng ức chế giúp khả năng kháng endolysin này ở chủng Staphylococcus aureus ( S. aureus ) Newman được ghi nhận tăng 50 . Trong một nghiên cứu khác, Sundarrajan và cộng sự đã báo cáo về khả năng kháng của các chủng thuộc chi Staphylococcus đối với P128, enzyme kháng khuẩn lai giữa EAD của endolysin thực khuẩn thể K và CBD của lysostaphin 51 . Tuy nhiên, các kiểu hình kháng phân lập được đều cho thấy tính nhạy cảm đối với kháng sinh β-lactam, suy giảm tốc độ tăng trưởng trong đánh giá in vitro và giảm độc lực trong đánh giá in vivo .
Những thuận lợi và khó khăn trong việc ứng dụng endolysin như một chất kháng khuẩn
Ngoài những ưu điểm về tính an toàn hay khả năng hình thành tính kháng đã được phân tích ở trên. Endolysin khi được sử dụng như một chất kháng khuẩn còn mang những ưu điểm như có thể tấn công tế bào vi khuẩn từ bên ngoài và ly giải tức thì sau khi tiếp xúc 52 , 53 ; có thể sử dụng đồng thời mà không bị giảm hoạt tính hoặc làm giảm hoạt tính của loại chất kháng khuẩn khác 54 , 55 , 56 ; khi sản xuất dưới dạng tái tổ hợp có thể sử dụng kỹ thuật gen điều chỉnh phổ kháng khuẩn, từ đó giúp hạn chế ảnh hưởng của endolysin đến hệ vi sinh vật trong môi trường sử dụng 57 ; một ưu điểm nổi trội hơn là endolysin đã được chứng minh vẫn giữ khả năng kháng khuẩn hiệu quả trong vùng nhiễm khuẩn đã hình thành biofilm hay các bề mặt mang màng nhầy khác 58 , 59 , 60 , 61 .
Các thách thức của việc ứng dụng endolysin chủ yếu đến từ bản chất là protein của loại phân tử sinh học này. Ngoài vấn đề giảm hiệu quả kháng khuẩn do gặp các kháng thể trung hòa khi đưa vào cơ thể, các nhà nghiên cứu cũng đang tập trung cải thiện tính bền cho các chế phẩm chứa endolysin, đặc biệt là các endolysin được sử dụng qua đường uống vì phân tử này cần đi qua môi trường khắc nghiệt của đường ruột. Một thách thức khác bắt nguồn từ việc tế bào vi khuẩn Gram âm tồn tại một màng đôi phospholipid bao bọc bên ngoài lớp peptidoglycan, điều này ngăn cản endolysin tiếp cận mục tiêu ly giải của mình. Vì lý do này, số lượng endolysin kháng vi khuẩn gây bệnh Gram âm đã được đưa vào ứng dụng vẫn còn hạn chế. Ngoài các endolysin tự nhiên cho thấy khả năng ly giải vi khuẩn Gram âm như LysPA26 kháng Pseudomonas aeruginosa ( P. aeruginosa ) 61 , Ply6A3 kháng Acinetobacter baumannii ( A.baumannii ) 62 , LysAB54 kháng nhiều chủng Gram âm 63 và một số endolysin khác, các endolysin còn lại để được sử dụng trong kháng vi khuẩn Gram âm cần được sử dụng cùng với một chất hỗ trợ hoặc đóng gói trong một hệ thống vận chuyển giúp endolysin thẩm thấu qua màng, hay phải được biến đổi di truyền để dung hợp với một nhân tố khác giúp endolysin có khả năng xuyên màng ngoài và tiếp cận được với lớp peptidoglycan 64 . Tuy nhiên, việc sử dụng các chiến lược này tạo ra thách thức khác cho các nhà nghiên cứu như lo ngại về tính an toàn của các chất hỗ trợ xuyên màng hay tốn thời gian và chi phí để nghiên cứu được hệ thống vận chuyển hiệu quả.
ỨNG DỤNG CỦA ENDOLYSIN
Ứng dụng trong điều trị bệnh trên người
Sự phổ biến của tình trạng kháng kháng sinh cùng với sự suy giảm tính hiệu quả của kháng sinh truyền thống là nguyên nhân dẫn đến việc các bệnh nhiễm khuẩn trên người ngày càng khó điều trị. Điều này thúc đẩy việc phát triển endolysin để trở thành một sản phẩm sinh dược, được sử dụng rộng rãi trong điều trị các bệnh nhiễm khuẩn ngoài da lẫn nhiễm khuẩn bên trong cơ thể ( Table 1 ). S aureus là một trong những vi khuẩn gây bệnh trên da phổ biến và nguy hiểm nhất bởi khả năng gây nhiễm khuẩn da và mô, lẫn độc lực cao của chúng khi xâm nhiễm vào máu, xương hoặc tim 65 . Quan trọng hơn, khả năng hình thành các chủng S. aureus kháng đa thuốc hoặc kháng methicillin (MRSA) khiến loài vi khuẩn này được ưu tiên phát triển các sản phẩm điều trị. Staphefekt SA.100 và Staphefekt XDR.300 là hai sản phẩm bôi ngoài da có thành phần là endolysin tái tổ hợp kháng S. aureus đã có mặt trên thị trường và dễ dàng tiếp cận đối với người bệnh. Bệnh cạnh đó, S. aureus còn là tác nhân dễ dàng lây nhiễm qua đường hô hấp, gây ra khoảng 30% các trường hợp nhiễm khuẩn đường hô hấp ở người trên toàn thế giới 66 . SAL200, sản phẩm đầu tiên có thành phần là endolysin được phát triển để điều trị nhiễm khuẩn MRSA. Khi được thử nghiệm sử dụng trực tiếp qua đường mũi trên mô hình chuột, sản phẩm này đã được chứng minh mang khả năng hỗ trợ điều trị bệnh viêm phổi do S. aureus gây ra 67 .
Bên cạnh các vi khuẩn Gram dương, một số loài gây bệnh Gram âm có độc lực cao và dễ sinh khả năng kháng thuốc như A. baumannii hay P. aeruginosa cũng là những tác nhân được tập trung nghiên cứu các sản phẩm điều trị. Tuy nhiên, như đã đề cập, lớp màng đôi bên ngoài của tế bào khiến việc phát triển các endolysin tự nhiên trở thành chất diệt vi khuẩn Gram âm gặp nhiều thách thức. Artilysin, một loại endolysin được biến đổi cấu trúc với khả năng xâm nhập màng ngoài tế bào vi khuẩn, là một trong những giải pháp hiệu quả đã được các nhà khoa học đề ra. Vào năm 2014, nhóm nghiên cứu của Briers đã giới thiệu artilysin LoGT-008 mang khả năng tiêu diệt cả P. aeruginosa và A. baumannii trong các thử nghiệm in vitro (thực hiện trên dòng tế bào sừng ở người) và in vivo (thực hiện trên mô hình Caenorhabditis elegans xâm nhiễm ruột) 68 .
Một bước tiến lớn gần đây trong việc ứng dụng rộng rãi các sản phẩm từ endolysin trong điều trị bệnh trên người là sự hợp tác giữa tập đoàn mỹ phẩm đa quốc gia L’Oréal và công ty Micreos, nơi phát triển endolysin XZ.700 kháng S. aureus điều trị bệnh viêm da cơ địa ở người 69 . Sự hợp tác này cho thấy tiềm năng phát triển lớn của endolysin để trở thành loại thuốc kháng khuẩn dễ dàng tiếp cận đối với tất cả mọi người.
Ứng dụng trong an toàn thực phẩm
Thực phẩm nhiễm khuẩn là nguy cơ gây tổn hại lớn nhất đối với ngành công nghiệp thực phẩm. Các tác nhân gây bệnh thường xuất hiện trong thực phẩm bao gồm S. aureus , chi Salmonella , Escherichia coli , L. monocytogenes , chi Clostridium ,… đã nhiều lần gây tổn hại to lớn đến sức khỏe và kinh tế. Cho đến hiện nay, đã có nhiều nghiên cứu thành công trong việc ứng dụng endolysin để kiểm soát tình trạng nhiễm khuẩn ở thực phẩm ( Table 2 ). Ví dụ tiêu biểu nhất là việc sử dụng endolysin như một chất bảo quản thực phẩm. Endolysin LysH5 đã được Obeso và các cộng sự chứng minh khả năng tiêu diệt S. aureus trong sữa thanh trùng và làm giảm mật độ vi khuẩn trong sữa xuống dưới ngưỡng phát hiện trong vòng bốn giờ 76 . Bên cạnh đó, các endolysin Ply118, Ply511 và PlyP35 từ thực khuẩn thể chi Listeria cũng được chứng minh khả năng diệt khuẩn cả trong điều kiện thường lẫn trong điều kiện nhiệt độ cao (90°C) 77 . Một số endolysin khác cũng mang tiềm năng ứng dụng trong bảo quản thực phẩm đã được công bố gần đây là endolysin từ thực khuẩn thể DLn1 với phổ kháng rộng được ứng dụng trong kiểm soát nhiễm khuẩn Bacillus cereus (B . cereus ) trong sữa 78 ; endolysin cpp-lys với tính bền được giữ vững trong khoảng nhiệt độ và pH rộng mang tiềm năng ứng dụng trong bảo quản lá xà lách khỏi các chủng vi khuẩn Clostridium perfringens ( C . perfringens ) tiết độc tố 79 ; endolysin LysPB32 được sử dụng kết hợp với các hợp chất từ tinh dầu thực vật giúp kiểm soát nhiễm khuẩn Salmonella Typhimurium trong các sản phẩm chứa thịt bò đã nấu chín 80 ; endolysin LysZC1 với độ bền cao mang khả năng kháng nhiều chủng vi khuẩn Gram âm là tác nhân nhiễm khuẩn thực phẩm 81 . Tại Việt Nam, nghiên cứu ứng dụng endolysin tiêu biểu nhất được tiến hành bởi nhóm nghiên cứu thuộc Viện Di truyền nông nghiệp với mục tiêu sản xuất endolysin LysSA29. Kết quả cho thấy khả năng kháng hiệu quả hai chủng gây bệnh S. aureus và Listeria monocytogenes ( L. monocytogenes ), đồng thời cho thấy tiềm năng cao trong việc bảo quản sữa tươi nguyên liệu và sữa thanh trùng 82 , 83 .
Bên cạnh khả năng bảo quản thực phẩm, endolysin còn được ứng dụng trong việc phát hiện thực phẩm nhiễm khuẩn dựa vào khả năng nhận diện đặc hiệu của CBD. Nhóm nghiên cứu của Kretzer và cộng sự đã chế tạo thành công hạt sắt từ được bọc CBD tái tổ hợp mang khả năng phát hiện L. monocytogenes với tỉ lệ phát hiện lên đến hơn 90%, cùng với đó là khả năng bắt giữ và phân tách vi khuẩn ra khỏi mẫu nhiễm 84 .
Ứng dụng trong nông nghiệp
Endolysin được ứng dụng rộng rãi trong việc bảo vệ các giống cây trồng và vật nuôi khỏi các vi khuẩn gây bệnh, từ đó giúp hạn chế lượng kháng sinh được sử dụng trong nông nghiệp ( Table 2 ). Ứng dụng tiêu biểu của endolysin trong trồng trọt đến từ việc tạo ra các giống cây trồng mang gene quy định endolysin, từ đó giúp cây trồng mang khả năng kháng đặc hiệu các loài vi khuẩn gây bệnh. Düring và cộng sự đã phát triển một giống khoai tây mới mang khả năng tổng hợp lysozyme từ thực khuẩn thể T4, giúp giống cây này có thể kháng lại vi khuẩn Erwinia carotovora gây thối rễ 85 . Một ý tưởng với sự kết hợp giữa endolysin và cây trồng đã được thực hiện bởi nhóm nghiên cứu của Oey. Trong nghiên cứu này, các tác giả đã sử dụng lục lạp trên cây thuốc lá làm “nhà máy” sản xuất các endolysin Cpl-1, Pal ( Streptococcus pneumoniae ) và PlyGBS (nhóm streptococcus) với sản lượng đạt 70% trên tổng lượng protein tan được cây tạo ra 86 , 87 . Bên cạnh việc tạo các giống cây trồng mang khả năng biểu hiện endolysin, enzyme này còn được sử dụng như một chất kháng khuẩn bảo vệ cây ăn trái giúp đảm bảo sản lượng. Endolysin từ thực khuẩn thể Otagovirus Psa374, kháng vi khuẩn Pseudomonas syringae pv. actinidiae gây bệnh trên cây kiwi, được sử dụng kết hợp với acid citric giúp làm giảm nồng độ vi khuẩn xuống dưới giới hạn phát hiện 88 .
Trong nông nghiệp chăn nuôi, khả năng kháng khuẩn của endolysin cũng được ứng dụng rộng rãi. Bệnh viêm vú do S. aureus là một trong nhưng tác nhân gây ảnh hưởng xấu đến ngành chăn nuôi gia súc. Protein λSA2E-LysK-SH3b, một enzyme tái tổ hợp giữa lysostaphin và endolysin đã được nghiên cứu và chứng minh hiệu quả kháng S. aureus trên mô hình chuột. Kết quả trên gợi ý rằng protein tái tổ hợp này có thể được sử dụng để điều trị bệnh viêm vú, từ đó ngăn chặn sự lây nhiễm của vi khuẩn này giữa các cá thể gia súc và từ cơ thể gia súc vào thực phẩm như sữa và thịt 89 .
Kết luận
Đa số các nghiên cứu sử dụng endolysin như hợp chất kháng khuẩn đã cho thấy tiềm năng ứng dụng của loại enzyme này. Mặc dù vậy, vẫn còn những thách thức trước mắt cần được giải quyết để phát triển endolysin trở thành các sản phẩm sinh dược mang hiệu quả điều trị cao. Các thách thức có thể được kể đến là khả năng hình thành tính kháng trên các loài vi khuẩn mục tiêu, thời gian bán hủy in vivo ngắn do sự sản sinh của các kháng thể trung hòa, hiệu quả diệt khuẩn không cao đặc biệt đối với các vi khuẩn Gram âm, và quan trọng hơn hết, là tính an toàn khi sử dụng endolysin trên người và động vật. Việc ngày càng nhiều các sản phẩm từ endolysin được phát triển và đưa ra thị trường chính là một tín hiệu tích cực trong công cuộc đẩy lùi tình trạng kháng kháng sinh với sự tham gia của loại enzyme này.
Danh mục từ viết tắt
A. baumannii : Acinetobacter baumannii ;
CBD: Cell-wall binding domain;
DNA: Deoxyribonucleic acid;
EAD: Enzymatically active domain;
GlcNAc: N-acetylglucosamine;
IgE: Immunoglobulin E;
IgG: Immunoglobulin G;
L. monocytogenes : Listeria monocytogenes ;
MRSA: Methicillin-resistant Staphylococcus aureus ;
MurNAc: N-acetylmuramic acid;
P. aeruginosa : Pseudomonas aeruginosa ;
S. aureus : Staphylococcus aureus ;
WHO: World Health Organization.
Xung đột lợi ích
Các tác giả không có xung đột lợi ích nào.
Đóng góp của tác giả
Lê Nguyễn Yến Vy: tổng hợp tài liệu, viết và chỉnh sửa.
Trần Văn Hiếu: đề xuất ý tưởng, chỉnh sửa và chấp thuận bản thảo.
Các tác giả đồng ý với bản cuối cùng của bản thảo.
References
- Antibiotic resistance. World Health Organization website (Newsroom/Fact. 2020;:. Google Scholar
- Việt Nam xuất hiện siêu vi khuẩn kháng tất cả kháng sinh, bác sĩ bất lực. . ;:. Google Scholar
- 33000 people die every year due to infections with antibiotic-resistant bacteria European Centre for Disease Prevention and Control2018. . ;:. Google Scholar
- Antibiotic Resistance Threats in the United States (2019 AR Threats Report. U.S. Centers for Disease Control and Prevention. ;:. Google Scholar
- D Duin, DL Paterson. Multidrug-resistant bacteria in the community: An update. Infect Dis Clin North Am. ;34(4):. Google Scholar
- MacNair C.R., Rutherford S.T., Tan M.-W.. Alternative therapeutic strategies to treat antibiotic-resistant pathogens. Nature Reviews Microbiology. 2024;22(5):. Google Scholar
- Golnari M., Bahrami N., Milanian Z., Rabbani Khorasgani M., Asadollahi M.A., Shafiei R.. Isolation and characterization of novel Bacillus strains with superior probiotic potential: comparative analysis and safety evaluation. Scientific Reports. ;14(1):. Google Scholar
- Leistikow K.R., Beattie R.E., Hristova K.R.. Probiotics beyond the farm: Benefits, costs, and considerations of using antibiotic alternatives in livestock. Frontiers in Antibiotics. 2021;1:. Google Scholar
- Abdelrahman F., Easwaran M., Daramola O.I., Ragab S., Lynch S., Oduselu T.J.. Phage-Encoded Endolysins. Antibiotics (Basel. 2021;10(2):. Google Scholar
- Nelson D.C., Schmelcher M., Rodriguez-Rubio L., Klumpp J., Pritchard D.G., Dong S.. Chapter 7 - Endolysins as Antimicrobials. Advances in Virus Research. 2012;:299-365. Google Scholar
- Gerstmans H., Criel B., Briers Y.. Synthetic biology of modular endolysins. Biotechnology Advances. 2018;36(3):624-40. Google Scholar
- Loessner M.J., Wendlinger G., Scherer S.. Heterogeneous endolysins in Listeria monocytogenes bacteriophages: a new class of enzymes and evidence for conserved holin genes within the siphoviral lysis cassettes. Mol Microbiol. 1995;16(6):. Google Scholar
- Schmelcher M., Shabarova T., Eugster M.R., Eichenseher F., Tchang V.S., Banz M.. Rapid multiplex detection and differentiation of Listeria cells by use of fluorescent phage endolysin cell wall binding domains. Appl Environ Microbiol. 2010;76(17):. Google Scholar
- Díaz E., López R., García J.L.. Chimeric phage-bacterial enzymes: a clue to the modular evolution of genes. Proc Natl Acad Sci U S A. 1990;87(20):. Google Scholar
- Schmelcher M., Donovan D.M., Loessner M.J.. Bacteriophage endolysins as novel antimicrobials. Future Microbiology. 2012;7(10):1147-71. Google Scholar
- Korndörfer I.P., Danzer J., Schmelcher M., Zimmer M., Skerra A., Loessner M.J.. The crystal structure of the bacteriophage PSA endolysin reveals a unique fold responsible for specific recognition of Listeria cell walls. J Mol Biol. 2006;364(4):. Google Scholar
- Cheng X., Zhang X., Pflugrath J.W., Studier F.W.. The structure of bacteriophage T7 lysozyme, a zinc amidase and an inhibitor of T7 RNA polymerase. Proc Natl Acad Sci U S A. 1994;91(9):. Google Scholar
- Briers Y., Volckaert G., Cornelissen A., Lagaert S., Michiels C., Hertveldt K.. Muralytic activity and modular structure of the endolysins of Pseudomonas aeruginosa bacteriophages φKZ and EL. Molecular Microbiology. 2007;65:. Google Scholar
- Pritchard D.G., Dong S., Kirk M.C., Cartee R.T., Baker JR. LambdaSa1 and LambdaSa2 prophage lysins of Streptococcus agalactiae. Appl Environ Microbiol. 2007;73(22):. Google Scholar
- García J.L., García E., Arrarás A., García P., Ronda C., López R.. Cloning, purification, and biochemical characterization of the pneumococcal bacteriophage Cp-1 lysin. J Virol. 1987;61(8):2573-80. Google Scholar
- Pritchard D.G., Dong S., Baker Engler, J.A. undefined. The bifunctional peptidoglycan lysin of Streptococcus agalactiae bacteriophage B30. Microbiology (Reading. 2004;150(Pt 7):. Google Scholar
- Becker S.C., Dong S., Baker Foster-Frey, J Pritchard, DG Donovan, D.M. undefined. LysK CHAP endopeptidase domain is required for lysis of live staphylococcal cells. FEMS Microbiol Lett. 2009;294(1):. Google Scholar
- Navarre W.W., Ton-That H., Faull K.F., Schneewind O.. Multiple enzymatic activities of the murein hydrolase from staphylococcal phage phi11. Identification of a D-alanyl-glycine endopeptidase activity. J Biol Chem. 1999;274(22):. Google Scholar
- Fischetti V.A.. Development of Phage Lysins as Novel Therapeutics: A Historical Perspective. Viruses. 2018;10(6):. Google Scholar
- Paradis-Bleau C., Cloutier I., Lemieux L., Sanschagrin F., Laroche J., Auger M.. Peptidoglycan lytic activity of the Pseudomonas aeruginosa phage phiKZ gp144 lytic transglycosylase. FEMS Microbiol Lett. 2007;266(2):. Google Scholar
- Lai M.J., Lin N.T., Hu A., Soo P.C., Chen L.K., Chen L.H.. Antibacterial activity of Acinetobacter baumannii phage φAB2 endolysin (LysAB2) against both gram-positive and gram-negative bacteria. Appl Microbiol Biotechnol. 2011;90(2):. Google Scholar
- Loessner M.J., Kramer K., Ebel F., Scherer S.. C-terminal domains of Listeria monocytogenes bacteriophage murein hydrolases determine specific recognition and high-affinity binding to bacterial cell wall carbohydrates. Mol Microbiol. 2002;44(2):. Google Scholar
- Matamp N., Bhat S.G.. Phage Endolysins as potential antimicrobials against multidrug resistant Vibrio alginolyticus and Vibrio parahaemolyticus: Current Status of Research and Challenges Ahead. Microorganisms. 2019;7(3):. Google Scholar
- Saier M.H., Reddy B.L.. Holins in Bacteria, Eukaryotes, and Archaea: Multifunctional xenologues with potential biotechnological and biomedical applications. Journal of Bacteriology. 2015;197(1):. Google Scholar
- Srividhya K.V., Krishnaswamy S.. Subclassification and targeted characterization of prophage-encoded two-component cell lysis cassette. J Biosci. 2007;32(5):. Google Scholar
- Berry J., Savva C., Holzenburg A., Young R.. The lambda spanin components Rz and Rz1 undergo tertiary and quaternary rearrangements upon complex formation. Protein Sci. 2010;19(10):. Google Scholar
- Twort F.. The transmissible bacteriallysin and its action on dead bacteria. The Lancet. 1925;206(5326):. Google Scholar
- Evans A.C.. Streptococcus Bacteriophage: A study of four serological types. Public Health Reports. 1934;1934;49(47):1386−401:. Google Scholar
- Maxted W.R.. The active agent in nascent phage lysis of streptococci. J Gen Microbiol. 1957;16(3):584-95. Google Scholar
- Reynals F.D.. Bacteriophage et microbes tues. Compt Rend Soc Biol. 1926;94:2. Google Scholar
- Gondil V.S., Harjai K., Chhibber S.. Endolysins as emerging alternative therapeutic agents to counter drug-resistant infections. Int J Antimicrob Agents. 2020;55(2):105844. Google Scholar
- M Loeffler Jutta, S Djurkovic, A Fischetti Vincent. Phage Lytic Enzyme Cpl-1 as a Novel Antimicrobial for Pneumococcal Bacteremia. Infection and Immunity. 2003;71(11):6199-204. Google Scholar
- Jun S.Y., Jung G.M., Yoon S.J., Oh M.-D., Choi Y.-J., Lee W.J.. Antibacterial properties of a pre-formulated recombinant phage endolysin, SAL-1. International Journal of Antimicrobial Agents. 2013;41(2):156-61. Google Scholar
- Grandgirard D., Loeffler J.M., Fischetti V.A., Leib S.L.. Phage lytic enzyme Cpl-1 for antibacterial therapy in experimental pneumococcal meningitis. J Infect Dis. 2008;197(11):1519-22. Google Scholar
- Jado I., López R., García E., Fenoll A., Casal J., García P.. Phage lytic enzymes as therapy for antibiotic-resistant Streptococcus pneumoniae infection in a murine sepsis model. J Antimicrob Chemother. 2003;52(6):967-73. Google Scholar
- Cheng Q., Nelson D., Zhu S., Fischetti V.A.. Removal of group B streptococci colonizing the vagina and oropharynx of mice with a bacteriophage lytic enzyme. Antimicrob Agents Chemother. 2005;49(1):111-7. Google Scholar
- Rashel M., Uchiyama J., Ujihara T., Uehara Y., Kuramoto S., Sugihara S.. Efficient elimination of multidrug-resistant Staphylococcus aureus by cloned lysin derived from bacteriophage phi MR11. J Infect Dis. 2007;196(8):1237-47. Google Scholar
- Fenton M., Casey P.G., Hill C., Gahan C.G., Ross R.P., McAuliffe O.. The truncated phage lysin CHAP(k) eliminates Staphylococcus aureus in the nares of mice. Bioeng Bugs. 2010;1(6):404-7. Google Scholar
- Abdelkader K., Gerstmans H., Saafan A., Dishisha T., Briers Y.. The Preclinical and clinical progress of bacteriophages and their lytic enzymes: The parts are easier than the whole. Viruses. 2019;11(2):96. Google Scholar
- Witzenrath M., Schmeck B., Doehn J.M., Tschernig T., Zahlten J., Loeffler J.M.. Systemic use of the endolysin Cpl-1 rescues mice with fatal pneumococcal pneumonia*. Critical Care Medicine. 2009;37(2):642-9. Google Scholar
- Zhang L., Li D., Li X., Hu L., Cheng M., Xia F.. LysGH15 kills Staphylococcus aureus without being affected by the humoral immune response or inducing inflammation. Sci Rep. 2016;6(29344):29344. Google Scholar
- Fischetti V.A.. Bacteriophage lytic enzymes: novel anti-infectives. Trends Microbiol. 2005;13(10):491-6. Google Scholar
- Jun S.Y., Jang I.J., Yoon S., Jang K., Yu K.-S., Cho J.Y.. Pharmacokinetics and Tolerance of the phage endolysin-based candidate drug SAL200 after a single intravenous administration among healthy volunteers:10.1128/aac.02629-16. Antimicrobial Agents and Chemotherapy. 2017;61(6):. Google Scholar
- Jun S.Y., Jung G.M., Yoon S.J., Youm S.Y., Han H.Y., Lee J.H.. Pharmacokinetics of the phage endolysin-based candidate drug SAL200 in monkeys and its appropriate intravenous dosing period. Clin Exp Pharmacol Physiol. 2016;43(10):1013-6. Google Scholar
- Becker S.C., Roach D.R., Chauhan V.S., Shen Y., Foster-Frey J., Powell A.M.. Triple-acting lytic enzyme treatment of drug-resistant and intracellular Staphylococcus aureus. Scientific Reports. 2016;6(1):25063. Google Scholar
- Sundarrajan S., Raghupatil J., Vipra A., Narasimhaswamy N., Saravanan S., Appaiah C.. Bacteriophage-derived CHAP domain protein, P128, kills Staphylococcus cells by cleaving interpeptide cross-bridge of peptidoglycan. Microbiology. 2014;160(10):2157-69. Google Scholar
- Oliveira H., Azeredo J., Lavigne R., Kluskens L.D.. Bacteriophage endolysins as a response to emerging foodborne pathogens. Trends in Food Science & Technology. 2012;28(2):103-15. Google Scholar
- Jeong A.L., Hakdong S., Sunggi H.. Exogenous Lytic Activity of SPN9CC Endolysin Against Gram-Negative Bacteria. J Microbiol Biotechnol. 2014;24(6):803-11. Google Scholar
- Ning H.-Q., Lin H., Wang J.-X.. Synergistic effects of endolysin Lysqdvp001 and ε-poly-lysine in controlling Vibrio parahaemolyticus and its biofilms. International Journal of Food Microbiology. 2021;343:109112. Google Scholar
- Hong H.W., Kim Y.D., Jang J., Kim M.S., Song M., Myung H.. Combination Effect of Engineered Endolysin EC340 With Antibiotics. Front Microbiol. 2022;13:. Google Scholar
- Zhang H., Zhang X., Liang S., Wang J., Zhu Y., Zhang W.. Bactericidal synergism between phage endolysin Ply2660 and cathelicidin LL-37 against vancomycin-resistant Enterococcus faecalis biofilms. npj Biofilms and Microbiomes. 2023;9(1):16. Google Scholar
- Kashani H.H., Schmelcher M., Sabzalipoor H., Hosseini E.S., Moniri R.. Recombinant Endolysins as Potential therapeutics against antibiotic-resistant Staphylococcus aureus: Current Status of Research and Novel Delivery Strategies. Clinical Microbiology Reviews. 2018;31(1):. Google Scholar
- Kuiper J.W.P., Hogervorst J.M.A., Herpers B.L., Bakker A.D., Klein-Nulend J., Nolte P.A.. The novel endolysin XZ.700 effectively treats MRSA biofilms in two biofilm models without showing toxicity on human bone cells in vitro. Biofouling. 2021;37(2):184-93. Google Scholar
- Cha Y., Son B., Ryu S.. Effective removal of Staphylococcal biofilms on various food contact surfaces by Staphylococcus aureus phage endolysin LysCSA13. Food Microbiol. 2019;84:. Google Scholar
- Yang D., Chen Y., Sun E., Hua L., Peng Z., Wu B.. Characterization of a lytic bacteriophage vB_EfaS_PHB08 harboring endolysin Lys08 against Enterococcus faecalis biofilms. Microorganisms. 2020;8(9):1332. Google Scholar
- Guo M., Feng C., Ren J., Zhuang X., Zhang Y., Zhu Y., Endolysin A.Novel Antimicrobial. LysPA26, against Pseudomonas aeruginosa. Front Microbiol. 2017;8(293):. Google Scholar
- Wu M., Hu K., Xie Y., Liu Y., Mu D., Guo H.. A Novel Phage PD-6A3, and Its Endolysin Ply6A3, With extended lytic activity against Acinetobacter baumannii. Front Microbiol. 2018;9(3302):. Google Scholar
- Khan F.M., Gondil V.S., Li C., Jiang M., Li J., Yu J.. A novel acinetobacter baumannii bacteriophage endolysin LysAB54 with high antibacterial activity against multiple Gram-negative microbes. front cell infect Microbiol. 2021;11:. Google Scholar
- Khan F.M., Rasheed F., Yang Y., Liu B., Zhang R.. Endolysins: a new antimicrobial agent against antimicrobial resistance. Strategies and opportunities in overcoming the challenges of endolysins against Gram-negative bacteria. Frontiers in Pharmacology. 2024;15:. Google Scholar
- Dryden M.S.. Complicated skin and soft tissue infection. Journal of Antimicrobial Chemotherapy. 2010;65(suppl_3):iii35-iii44. Google Scholar
- Wertheim H.F.L., Melles D.C., Vos M.C., Leeuwen W., Belkum A., Verbrugh H.A.. The role of nasal carriage in Staphylococcus aureus infections. The Lancet Infectious Diseases. 2005;5(12):751-62. Google Scholar
- Bae J.Y., Jun K.I., Kang C.K., Song K.H., Choe P.G., Bang J.H.. Efficacy of Intranasal Administration of the Recombinant Endolysin SAL200 in a Lethal Murine Staphylococcus aureus pneumonia model. Antimicrob Agents Chemother. 2019;63(4):. Google Scholar
- Briers Y., Walmagh M., Puyenbroeck Cornelissen, A Cenens, W. undefined. Aertsen AEngineered Endolysin-Based “Artilysins. To Combat Multidrug-Resistant Gram-Negative Pathogens. mBio. 2014;5(4):. Google Scholar
- Murray E., Draper L.A., Ross R.P., Hill C.. The advantages and challenges of using endolysins in a clinical setting. Viruses. 2021;13(4):. Google Scholar
- Totté J.E.E., Doorn M.B., Pasmans S.. Successful Treatment of Chronic Staphylococcus aureus-Related Dermatoses with the Topical Endolysin Staphefekt SA.100: A Report of 3 Cases. Case Rep Dermatol. 2017;9(2):19-25. Google Scholar
- Pallesen E.M.H., Gluud M., Vadivel C.K., Buus T.B., Rooij B., Zeng Z.. Endolysin Inhibits skin colonization by Patient-derived Staphylococcus aureus and Malignant T-Cell activation in cutaneous T-cell lymphoma. Journal of Investigative Dermatology. 2023;143(9):1757-68. Google Scholar
- Danis-Wlodarczyk K.M., Wozniak D.J., Abedon S.T.. Treating bacterial infections with bacteriophage-Based enzybiotics: in vitro. In vivo and clinical application. 2021;10 (12):. Google Scholar
- Schuch R., Lee H.M., Schneider B.C., Sauve K.L., Law C., Khan B.K.. Combination Therapy With Lysin CF-301 and Antibiotic Is Superior to Antibiotic Alone for Treating Methicillin-Resistant Staphylococcus aureus–Induced Murine Bacteremia. The Journal of Infectious Diseases. 2013;209(9):1469-78. Google Scholar
- Indiani C., Sauve K., Raz A., Abdelhady W., Xiong Y.Q., Cassino C., Lysin C.F.-301 The Antistaphylococcal. Activates key host factors in human blood to potentiate methicillin-resistant Staphylococcus aureus bacteriolysis. Antimicrob Agents Chemother. 2019;63(4):. Google Scholar
- Channabasappa S., Durgaiah M., Chikkamadaiah R., Kumar S., Joshi A., Sriram B.. Efficacy of novel antistaphylococcal ectolysin P128 in a rat model of methicillin-resistant Staphylococcus aureus bacteremia. Antimicrob Agents Chemother. 2018;62(2):. Google Scholar
- Obeso J.M., Martínez B., Rodríguez A., García P.. Lytic activity of the recombinant staphylococcal bacteriophage PhiH5 endolysin active against Staphylococcus aureus in milk. Int J Food Microbiol. 2008;128(2):212-8. Google Scholar
- Schmelcher M., Waldherr F., Loessner M.J.. Listeria bacteriophage peptidoglycan hydrolases feature high thermoresistance and reveal increased activity after divalent metal cation substitution. Appl Microbiol Biotechnol. 2012;93(2):633-43. Google Scholar
- Li N., Yuan X., Li C., Chen N., Wang J., Chen B.. A novel Bacillus cereus bacteriophage DLn1 and its endolysin as biocontrol agents against Bacillus cereus in milk. International Journal of Food Microbiology. 2022;369(109615):. Google Scholar
- Zhao X., Li L., Zhang Q., Li M., Hu M., Luo Y.. Characterization of the Clostridium perfringens phage endolysin cpp-lys and its application on lettuce. International Journal of Food Microbiology. 2023;405(110343):. Google Scholar
- Kim J., Kim S., Wang J., Ahn J.. Synergistic antimicrobial activity of essential oils in combination with phage endolysin against Salmonella Typhimurium in cooked ground beef. Food Control. 2024;157(110187):. Google Scholar
- Abdelrahman F., Gangakhedkar R., Nair G., El-Didamony G., Askora A., Jain V.. Pseudomonas phage ZCPS1 endolysin as a potential therapeutic agent. Viruses. 2023;15(2):. Google Scholar
- NTG Nguyễn Thị Hồng Hải, Thu Phạm Thị Lý. Nghiên cứu sản xuất chế phẩm enzyme Endolysins LysSA29 từ chủng vi khuẩn tái tổ hợp quy mô 80. . ;:. Google Scholar
- Viện Khoa học và Công nghệ. Kiểm tra, thẩm định sản phẩm nhiệm vụ KHCN tại Viện Di truyền nông nghiệp 2019. . ;:. Google Scholar
- Kretzer J.W., Lehmann R., Schmelcher M., Banz M., Kim K.-P., Korn C.. Use of high-affinity cell wall-binding domains of bacteriophage endolysins for immobilization and separation of bacterial cells. Applied and Environmental Microbiology. 2007;73(6):1992-2000. Google Scholar
- Düring K., Porsch P., Fladung M., Lörz H.. Transgenic potato plants resistant to the phytopathogenic bacterium Erwinia carotovora. Plant Journal. 1993;3(587):587-98. Google Scholar
- Oey M., Lohse M., Scharff L.B., Kreikemeyer B., Bock R.. Plastid production of protein antibiotics against pneumonia via a new strategy for high-level expression of antimicrobial proteins. Proc Natl Acad Sci U S A. 2009;106(16):6579-84. Google Scholar
- Oey M., Lohse M., Kreikemeyer B., Bock R.. Exhaustion of the chloroplast protein synthesis capacity by massive expression of a highly stable protein antibiotic. Plant J. 2009;57(3):436-45. Google Scholar
- Sisson H.M., Fagerlund R.D., Jackson S.A., Briers Y., Warring S.L., Fineran P.C.. Antibacterial synergy between a phage endolysin and citric acid against the Gram-negative kiwifruit pathogen Pseudomonas syringae pv. actinidiae. Applied and Environmental Microbiology. ;90(3):. Google Scholar
- Schmelcher M., Powell A.M., Becker S.C., Camp M.J., Donovan D.M.. Chimeric phage lysins act synergistically with lysostaphin to kill mastitis-causing Staphylococcus aureus in murine mammary glands. Appl Environ Microbiol. 2012;78(7):2297-305. Google Scholar





