
 Open Access
Open Access Abstract
Eggplant (Solanum melongena L.), an important vegetable crop because of its nutritional and medicinal values, is often attacked by insects and pathogens. Quantitative gene expression analysis by reverse transcription‒quantitative PCR (RT‒qPCR) is an effective and common approach to analyze the function of genes involving in the stress responses in eggplant. The RT‒qPCR method always included the step of normalizing the expression data of the target gene to that of a stably expressed internal reference gene to ensure the accuracy of the quantitative results. This paper presented the evaluation of the transcriptional expression of adenine phosphoribosyl transferase (SmAPRT) gene in different tissues of S. melongena (young leaf, mature leaf, stem, root, flower, and fruit) under normal conditions. Besides, the expression of SmAPRT was also measured in leaf tissues of plants damaged by mechanical wounding, plants treated with stress signal molecules induced by methyl jasmonate (MeJA) or methyl salicylate (MeSA), and untreated control plants. PCR with a SmAPRT-specific primer pair (SmAPRT-qF as forward primer and SmAPRT-qR as reverse primer) amplified a 103-bp fragment from S. melongena with the efficiency of 92.7%. This value was within the recommended range (90‒110%). The Ct values of qPCR reactions using cDNA template from the wounded plants or plants treated with MeJA or MeSA were not statistically significantly different from the Ct ones of the control plants. This SmAPRT gene was also stably expressed in all tested eggplant tissues. Therefore, SmAPRT would be used as a reference gene to normalize expression data of genes of interest in different tissues as well as in the response of S. melongena to the specific stress conditions as stated above.
MỞ ĐẦU
Cây cà tím ( Solanum melongena ) là loài nông sản quan trọng và phổ biến ở các nước Châu Á, Trung Đông, Cận Đông, Địa Trung Hải và Châu Phi. Cà tím được sử dụng để điều trị một số bệnh như tiểu đường, viêm khớp, hen suyễn và viêm phế quản 1 . Vì hiệu quả biến nạp gene cao, cây cà tím còn được sử dụng làm cây mô hình để nghiên cứu các tính trạng nông học khác nhau thông qua công nghệ chuyển gene 2 . Hiện tại, cơ sở dữ liệu bộ gene cà tím đã được giải mã tương đối hoàn chỉnh, với khoảng 35.000 gene được chú thích 3 . Đây là một nguồn dữ liệu hữu ích để phân tích chức năng các gene mới, đặc biệt là các gene tham gia trong con đường truyền tín hiệu căng thẳng (stress) ở loài này 4 , 5 , 6 .
Cây cà tím là cây hằng niên, quả mọng và được thu hoạch liên tục, khiến cho việc bảo vệ cây khỏi nấm bệnh và sâu hại gặp nhiều khó khăn. Hơn nữa, do người tiêu dùng thường để nguyên vỏ quả khi chế biến cà tím, người trồng cần hạn chế sử dụng các loại thuốc bảo vệ thực vật trên loại cây này. Thực vật nói chung và cây cà tím nói riêng phản ứng với các tác nhân gây căng thẳng bằng cách cảm ứng các con đường dẫn truyền tín hiệu, thông qua việc kích hoạt tổng hợp các hormone thực vật hoạt động như những tín hiệu phòng vệ bao gồm jasmonic acid (JA), salicylic acid (SA) và ethylene (ET) 7 , 8 , dẫn đến những thay đổi trong sự biểu hiện của các gene liên quan đến cơ chế phòng vệ 7 , 9 . Việc phân tích các tác động của tổn thương cơ học và tác động của tín hiệu gây căng thẳng (JA/SA) đến sự biểu hiện của các gene liên quan đến cơ chế phòng vệ có thể góp phần làm sáng tỏ cơ chế phân tử của những quá trình này.
Ngày nay, phương pháp định lượng RT-qPCR đã trở thành một trong những phương pháp được sử dụng rộng rãi để định lượng nhanh chóng và chính xác sự biểu hiện của gene ở mức phiên mã. Tuy nhiên, sự chính xác của phương pháp định lượng RT-qPCR phụ thuộc vào việc chọn gene tham chiếu (reference genes) phù hợp để có thể chuẩn hóa dữ liệu biểu hiện của các gene mục tiêu. Việc sử dụng gene tham chiếu giúp kiểm soát sự khác biệt về số lượng và chất lượng RNA tách chiết giữa các mẫu khảo sát, hiệu suất của phản ứng phiên mã ngược, tính đặc hiệu của cặp mồi và hiệu suất của quá trình nhân bản. Một số gene giữ nhà (housekeeping genes) liên quan đến các hoạt động thiết yếu của tế bào như các gene mã hóa 18S rRNA, 25S rRNA, glyceraldehyde-3-phosphate dehydrogenase (GAPDH) và ubiquitin (UBQ) thường được sử dụng làm gene tham chiếu trong các phân tích RT-qPCR vì chúng có mức biểu hiện ổn định trong các điều kiện thí nghiệm khác nhau 10 , 11 , 12 , 13 , 14 . Tuy nhiên, các nghiên cứu gần đây chỉ ra rằng sự biểu hiện của các gene giữ nhà cũng thay đổi đáng kể trong nhiều trường hợp 15 , 16 , 17 . Do đó, trước khi phân tích sự biểu hiện phiên mã của gene mục tiêu trên một hệ thống sinh học cụ thể, cần đánh giá tính ổn định của gene tham chiếu được chọn trên chính hệ thống đó 18 , 19 .
Sự biểu hiện phiên mã của các gene 18S rRNA, adenine phosphoribosyl transferase (APRT), GAPDH, cyclophilin, actin, và ribulose-1,5-bisphosphate carboxylase (RuBP) đã được khảo sát trên 6 mô khác nhau bao gồm lá non, lá trưởng thành, chồi, rễ, nụ hoa và hoa 20 . Kết quả phân tích dữ liệu biểu hiện gene bằng ba thuật toán thống kê khác nhau (geNorm, NormFinder và BestKeeper) đã cho thấy rằng 18S rRNA, cyclophilin và APRT là các gene có sự biểu hiện ổn định nhất ở các mô nêu trên 21 . Kết quả này được ghi nhận trên 4 giống cà tím khác nhau (Pusa Shyamla, Pusa Ankur, Pusa Kranti và Pusa Uttam), 2 dòng cà tím chuyển gene và 2 dòng cà tím bị nhiễm tuyến trùng.
Bài báo này trình bày khả năng sử dụng APRT làm gene tham chiếu trong phân tích biểu hiện gene bằng phương pháp RT-qPCR ở cà tím, khi có tổn thương cơ học hoặc dưới ảnh hưởng của xử lý với MeJA hoặc MeSA. Tính ổn định trong sự biểu hiện của gene SmAPRT cũng được đánh giá trên các mô khác nhau của cây cà tím, bao gồm những mô đã được khảo sát trước đây gồm lá non, lá trưởng thành, rễ, và hoa, cùng những mô chưa được khảo sát như thân và quả.
VẬT LIỆU VÀ PHƯƠNG PHÁP
Vật liệu nghiên cứu
Hạt cà tím ( Solanum melongena L.) dòng thuần (giống cà tím Mê Linh quả dài) được cung cấp bởi Viện Cây ăn quả miền Nam (SOFRI). Hạt được gieo trồng trong nhà màng của Phòng thí nghiệm Sinh hóa Ứng dụng, Bộ môn Sinh hóa và được định danh tại Bộ môn Sinh thái-Sinh học tiến hóa, Khoa Sinh học‒Công Nghệ Sinh Học, Trường Đại học Khoa học Tự nhiên, Đại học Quốc gia Tp. HCM.
Thu mẫu mô của cà tím
Chín cây cà tím 13 tuần tuổi ( Figure 1 ) có kích thước đồng đều nhau được sử dụng để thu mẫu lá non, lá trưởng thành, thân, rễ, hoa và quả. Mỗi nghiệm thức bao gồm 3 tổ hợp, với mỗi tổ hợp bao gồm 3 cây cà tím.
Xử lý tạo vết thương cơ học và xử lý với tín hiệu căng thẳng (MeJA/MeSA) trên cây cà tím
Ba mươi sáu cây cà tím 6 tuần tuổi, có kích thước đồng đều, không bị côn trùng và mầm bệnh tấn công được lựa chọn cho 4 nghiệm thức bao gồm xử lý tạo vết thương, xử lý với MeJA, xử lý với MeSA và đối chứng. Mỗi nghiệm thức có 3 tổ hợp, với mỗi tổ hợp bao gồm 3 cây cà tím.
Ở nghiệm thức xử lý tạo vết thương: một lá ở tầng trên cùng (ngay dưới chồi cây) bị gây tổn thương cơ học bằng cách dùng kẹp hemostate bấm 3 đường ngang qua gân chính và gân phụ của lá ( Figure 2 ). Sau 24 giờ xử lý, một lá nguyên vẹn ở ngay tầng dưới của lá bị gây tổn thương được thu nhận và bảo quản trong nitrogen lỏng.
Ở nghiệm thức xử lý với MeJA: mỗi cây được đặt trong một hộp thủy tinh kín có chứa một mẩu giấy lọc được tẩm 6,8 mg MeJA 22 ( Figure 2 ). Sau 2 giờ xử lý, cây được đặt trở lại điều kiện sinh dưỡng bình thường trước đó để tiếp tục tăng trưởng. Sau 24 giờ, mô lá của mỗi cây được thu nhận và bảo quản trong nitrogen lỏng.
Ở nghiệm thức xử lý với MeSA: thực hiện tương tự như nghiệm thức xử lý MeJA, với mẩu giấy lọc được tẩm 3 mg MeSA 23 ( Figure 2 ).
Ở nghiệm thức đối chứng (control): cây không bị gây tạo vết thương và không trải qua xử lý MeJA hoặc MeSA.
Figure 1 . Cây cà tím 13 tuần tuổi được sử dụng để thu các mẫu mô lá non, lá trưởng thành, thân, rễ, hoa và quả

Figure 2 . Thí nghiệm mô phỏng cây cà tím chịu tác động của các tác nhân gây căng thẳng

Tách chiết RNA tổng số và tổng hợp cDNA
Mẫu mô cà tím từ mỗi tổ hợp được nghiền thành dạng bột trong nitrogen lỏng. RNA tổng số được tách chiết từ 50 mg bột mô bằng kít EZ-10 Spin column Plant RNA Mini-preps (Bio Basic, Canada). DNA bộ gene còn hiện diện trong mẫu RNA tách chiết được loại bỏ theo hướng dẫn của kít RapidOut DNA Removal (Thermo Scientific, USA). Mẫu RNA có chất lượng tốt thường hiển thị rõ 2 vạch 28S và 18S khi điện di trên gel agarose 1%. Chất lượng RNA được đánh giá dựa trên tỷ lệ của sự hấp thụ tại bước sóng 260 và 280 nm (A 260 /A 280 ). Hàm lượng RNA được xác định bằng phương pháp đo mật độ quang trên máy quang phổ NanoVue Plus 7415 (Biochrom, UK). Quá trình tổng hợp cDNA được thực hiện theo hướng dẫn của kít Reverse transcriptase Aid First Strand (Thermo Fisher Scientific, USA), sử dụng 1,5 µg RNA cho phản ứng có thể tích 20 µL. cDNA được sử dụng làm khuôn cho các phản ứng qPCR.
Thiết kế cặp mồi nhân bản đặc hiệu một đoạn trình tự DNA từ bản sao của gene SmAPRT
Mồi xuôi SmAPRT-qF và mồi ngược SmAPRT-qR sử dụng trong phản ứng RT-qPCR được thiết kế dựa trên trình tự mã hóa APRT (Genbank: JX448345) 21 theo nguyên tắc sau: i / Chiều dài của mồi (bp), hàm lượng GC (%) và nhiệt độ nóng chảy (°C) trong khoảng khuyến nghị và các thông số này được kiểm tra bằng phần mềm OligoAnalyzer 3.1 20 ; ii/ Khoảng cách giữa mồi xuôi và mồi ngược dao động trong khoảng 75‒150 bp; iii/ Ít nhất một trong hai mồi bắt cặp bổ sung với vùng trình tự tiếp nối giữa hai exon kề nhau để mồi chỉ bắt cặp với SmAPRT cDNA nhưng không bắt cặp với trình tự gene mục tiêu (trong trường hợp DNA bộ gene còn lẫn trong mẫu RNA tách chiết); iv/ Cặp mồi bắt cặp đặc hiệu với một trình tự duy nhất là bản sao của gene SmAPRT .
Khảo sát nhiệt độ gắn mồi và kiểm tra tính đặc hiệu của mồi trong phản ứng qPCR
Phản ứng qPCR được thực hiện trên hệ thống real-time PCR CFX96-Touch system (BioRad, USA) tại 4 điểm nhiệt độ gắn mồi (T a ): 52 °C; 55 °C; 56,3 °C và 60 °C. Thành phần của phản ứng qPCR (tổng thể tích 12,5 μL) bao gồm: 6,7 μL nước đã loại nuclease; 1,25 μL đệm phản ứng h-Taq 10X; 0,6 μL MgCl 2 50 mM; 0,25 μL dNTP 10 mM; 0,5 μL mồi xuôi SmAPRT-qF 10 mM; 0,5 μL mồi ngược SmAPRT-qR 10 mM; 0,6 μL EvaGreen ® Dye 20X; 0,1 μL SolGent™ h-Taq DNA polymerase 2,5 U/μL; 2 μL cDNA. Chu trình nhiệt của phản ứng qPCR như sau: 95 °C/ 15 phút, 40 chu kỳ của 95 °C (20 giây), T a °C (40 giây) và 72 °C (6 giây), trong đó T a (annealing temperature) là nhiệt độ gắn mồi. Đường cong nóng chảy (melting curve) sau khi phản ứng qPCR kết thúc được thiết lập theo chương trình nhiệt độ như sau: tăng dần nhiệt độ từ 55 đến 95 °C, mỗi lần tăng 0,5 °C, giữ lại trong 5 giây và đọc tín hiệu. Nhiệt độ gắn mồi được chọn để thực hiện các khảo sát tiếp theo là nhiệt độ tại đó mẫu chứng âm cho kết quả âm tính (không ngoại nhiễm), còn mẫu khảo sát chỉ cho một đỉnh tín hiệu huỳnh quang duy nhất trong khoảng 75‒90 °C trên đường cong nóng chảy (ứng với sản phẩm nhân bản đặc hiệu); đồng thời, phản ứng tại nhiệt độ gắn mồi đó có giá trị chu kỳ ngưỡng C t thấp nhất. Để kiểm chứng rằng phản ứng qPCR với cặp mồi trên đã tạo ra sản phẩm nhân bản duy nhất và đặc hiệu, sản phẩm nhân bản được điện di trên gel agarose 1%, được tinh sạch và gửi giải trình tự hai chiều với mồi SmAPRT-qF và SmAPRT-qR tại 1 st Base (Malaysia).
Khảo sát hiệu suất nhân bản
Hiệu suất nhân bản của phản ứng qPCR với cặp mồi SmAPRT-qF và SmAPRT-qR tại nhiệt độ gắn mồi đã khảo sát được đánh giá thông qua đường chuẩn biểu diễn mối tương quan giữa chu kỳ ngưỡng (C t ) với số lượng bản sao trong các mẫu cDNA đã được pha loãng theo hệ số bậc 10. Hiệu suất nhân bản của phản ứng qPCR (E%) được tính toán theo công thức: E% = (n -1/a -1) × 100, với n là hệ số pha loãng cDNA và a là hệ số góc của đường chuẩn có dạng y = a x + b 24 , 25 . Phản ứng qPCR cho kết quả định lượng tin cậy khi hệ số tương quan R 2 ≥ 0,985 và hiệu suất nhân bản (E%) trong khoảng 90-110% 24 , 26 .
Khảo sát sự biểu hiện phiên mã của gene SmAPRT ở cà tím
Mức độ biểu hiện phiên mã của gene SmAPRT trong các mô khác nhau được khảo sát bằng phương pháp real-time PCR với các điều kiện phản ứng đã được thiết lập như trên. Khuôn được sử dụng trong phản ứng qPCR là các mẫu cDNA được tổng hợp từ RNA tổng số thu nhận từ mô lá non, lá trưởng thành, thân, rễ, hoa và quả của cây cà tím.
Tương tự, mức độ biểu hiện phiên mã của gene SmAPRT trong mô lá của các cây chịu tác động của các tác nhân gây căng thẳng bao gồm xử lý gây tạo vết thương cơ học, xử lý với MeJA và xử lý với MeSA cũng được khảo sát bằng phương pháp qPCR với các điều kiện phản ứng tương tự. Giá trị chu kỳ ngưỡng C t trong nghiệm thức đối chứng và xử lý được ghi nhận và so sánh bằng phân tích thống kê.
Xử lý thống kê
Phương pháp phân tích thống kê ANOVA một yếu tố (Tukey HSD test) được sử dụng để so sánh các giá trị chu kỳ ngưỡng C t trung bình (C t-tb ) của các phản ứng qPCR trong thí nghiệm khảo sát nhiệt độ gắn mồi, và trong thí nghiệm khảo sát biểu hiện của gene SmAPRT theo mô ở cây cà tím. Ở thí nghiệm mô phỏng cây cà tím chịu tác động của các tác nhân gây căng thẳng, phương pháp Student’s t -test (two-tail, unpaired, equal variance) được sử dụng để so sánh giá trị chu kỳ ngưỡng C t-tb của các phản ứng qPCR trong nghiệm thức xử lý với giá trị C t-tb trong nghiệm thức đối chứng.
KẾT QUẢ VÀ THẢO LUẬN
Thiết kế cặp mồi nhân bản đặc hiệu một đoạn trình tự DNA từ bản sao của gene SmAPRT
Trình tự, thông số và vị trí của mồi xuôi SmAPRT-qF và mồi ngược SmAPRT-qR được trình bày trong Table 1 và Figure 3 . Cặp mồi được dự đoán cho phép nhân bản đặc hiệu một đoạn trình tự DNA có chiều dài 103 bp.
Figure 3 . Vị trí bắt cặp của mồi xuôi SmAPRT-qF và mồi ngược SmAPRT-qR trên một phần trình tự mã hóa SmAPRT (Genbank: JX448345) . Các exon kề nhau được phân biệt với nhau bởi màu nền.

Tách chiết RNA tổng số
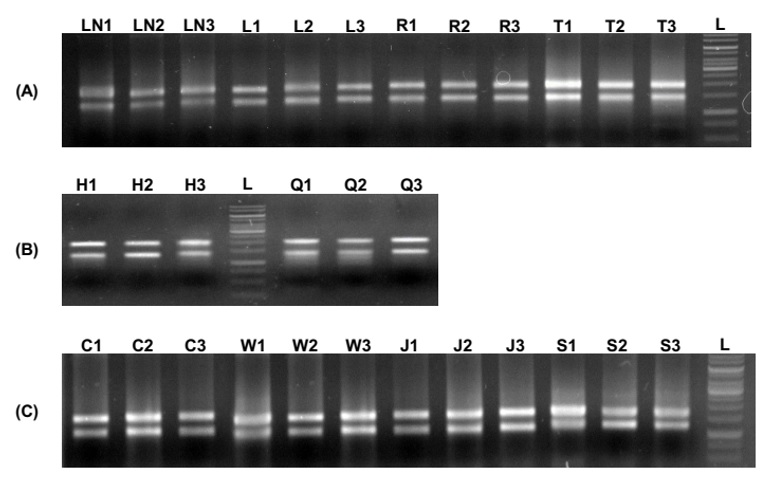
Các mẫu RNA tách chiết được điện di trên gel agarose 1%. Hình ảnh từ bản gel ( Figure 4 ) cho thấy các mẫu RNA đều có chất lượng tốt với các vạch 18S và 28S hiện diện rõ nét.
Figure 4 . Kết quả điện di mẫu RNA tổng số tách chiết từ các mô khác nhau (A và B) và từ mô lá trong thí nghiệm mô phỏng cây cà tím chịu tác động của các tác nhân gây căng thẳng (C). LN: lá non; L: lá trưởng thành; R: rễ; T: thân; H: hoa; Q: quả; C: đối chứng; W: xử lý gây tạo vết thương; J: xử lý với MeJA; S: xử lý với MeSA; L: thang chuẩn DNA 1kb.
Khảo sát nhiệt độ gắn mồi trong phản ứng qPCR
Mẫu cDNA của mô quả (pha loãng 10 lần) được sử dụng làm khuôn trong phản ứng. Kết quả ở Figure 5 cho thấy các đường cong nóng chảy ứng với các điểm nhiệt độ gắn mồi được khảo sát đều chỉ có một đỉnh tín hiệu huỳnh quang duy nhất trong khoảng nhiệt độ 81,5‒82,5 °C. Dựa trên phân tích thống kê ANOVA một yếu tố - Tukey HSD test, các giá trị chu kỳ ngưỡng C t-tb của các phản ứng qPCR sử dụng nhiệt độ gắn mồi khác nhau ( Table 2 ) không có sự khác biệt có ý nghĩa thống kê (p-value = 0,088 > 0,05) ( Table 5 ). Như vậy, trong các thí nghiệm tiếp theo, có thể sử dụng một trong 4 nhiệt độ gắn mồi nêu trên. Cụ thể, 55 °C đã được chọn làm nhiệt độ gắn mồi.
Figure 5 . Đường cong nóng chảy của sản phẩm nhân bản trong các phản ứng qPCR sử dụng các nhiệt độ gắn mồi (T a ) khác nhau

Phân tích trình tự của đoạn DNA được nhân bản trong phản ứng qPCR
Hình ảnh điện đi sản phẩm nhân bản của phản ứng qPCR sử dụng cặp mồi SmAPRT-qF và SmAPRT-qR cho thấy ở giếng M của bản gel xuất hiện một vạch có vị trí thấp hơn vạch 250 bp của thang chuẩn DNA (Thermo Fisher Scientific, USA). Vạch này phù hợp với kích thước dự đoán 103 bp của sản phẩm nhân bản đặc hiệu ( Figure 6 ). Bên cạnh đó, ở giếng đối chứng (sử dụng nước đã loại nuclease thay cho cDNA) không xuất hiện vạch có kích thước tương ứng ( Figure 6 ). Sản phẩm nhân bản sau đó được tinh sạch từ gel agarose và giải trình tự hai chiều với mồi SmAPRT-qF và SmAPRT-qR. Kết quả giải trình tự ( Figure 7 ) cho thấy đoạn DNA này chính là sản phẩm nhân bản duy nhất và đặc hiệu của phản ứng qPCR.
Figure 6 . Kết quả điện di sản phẩm qPCR sử dụng cặp mồi SmAPRT-qF và SmAPRT-qR trên gel agarose 1%. ĐC: Đối chứng âm không bổ sung cDNA; T: Thang chuẩn DNA 1kb; M: Sản phẩm nhân bản sử dụng khuôn cDNA chuẩn bị từ mô quả cà tím.

Figure 7 . Phân tích trình tự đoạn DNA được nhân bản trong phản ứng qPCR với cặp mồi SmAPRT-qF và SmAPRT-qR. (A) Kết quả giải trình tự sản phẩm qPCR với mồi xuôi SmAPRT-qF; (B) Kết quả giải trình tự sản phẩm qPCR với mồi ngược SmAPRT-qR; (C) Kết quả sắp gióng cột trình tự đoạn DNA được nhân bản theo cơ sở dữ liệu (Genbank: JX448345) và theo kết quả giải trình tự.

Hiệu suất nhân bản của phản ứng qPCR
Để đánh giá hiệu suất nhân bản của phản ứng qPCR, đường chuẩn biểu diễn mối tương quan giữa chu kỳ ngưỡng (C t ) với số lượng bản sao hiện diện trong mẫu cDNA đã pha loãng nồng độ theo hệ số 10 được xây dựng như mô tả trong Figure 8 . Đồ thị đường chuẩn có hệ số tương quan R 2 = 0,9996 và có phương trình y= -3,511x + 20,167; trong đó hệ số góc ( a ) = -3,511 ( Figure 8 ). Hiệu suất nhân bản của phản ứng qPCR được tính toán dựa trên hệ số góc của đường chuẩn là 92,7 %. Giá trị hiệu suất này nằm trong phạm vi khuyến nghị (90‒110 %) đảm bảo độ tin cậy của kết quả định lượng.
Figure 8 . Đồ thị đường chuẩn biểu diễn mối tương quan giữa chu kỳ ngưỡng (C t ) và nồng độ cDNA trong phản ứng qPCR

Sự biểu hiện phiên mã của gene SmAPRT ở các mô khác nhau của cà tím
Dựa trên các dữ liệu nghiên cứu so sánh độ ổn định trong biểu hiện phiên mã của các gene tham chiếu tiềm năng ở cây cà tím , SmAPRT được đánh giá là gene có sự biểu hiện phiên mã tương đối ổn định trong 6 mô đại diện cho các giai đoạn phát triển của cà tím (lá non, lá trưởng thành, chồi, rễ, nụ hoa và hoa) 21 . Trong nghiên cứu này, tính ổn định trong sự biểu hiện của gene SmAPRT được khảo sát một cách đầy đủ hơn trên các mô khác nhau của cây cà tím bao gồm cả những mô chưa được sử dụng trong nghiên cứu nêu trên như mô thân và mô quả. Giá trị C t của các phản ứng qPCR với cặp mồi SmAPRT-qF và SmAPRT-qR trên các mẫu cDNA lần lượt được chuẩn bị từ mô lá non, lá trưởng thành, thân, rễ, hoa và quả được thể hiện ở Table 3 . Giá trị chu kỳ ngưỡng (C t ) của SmAPRT ở các mô khác nhau không có sự khác biệt có ý nghĩa thống kê (p-value = 0,70 > 0,05) theo phân tích ANOVA một yếu tố ( Table 6 ), chứng tỏ gene này biểu hiện ổn định ở các mô cà tím được khảo sát. Kết quả này tương đồng với nhận định của Gantasala và cộng sự (2013) khi phân tích tính ổn định trong biểu hiện phiên mã của gene SmAPRT trên 6 mẫu mô đại diện 6 giai đoạn phát triển khác nhau của cà tím gồm: lá non, lá trưởng thành, chồi, rễ, nụ hoa và hoa. Đồng thời, kết quả của nghiên cứu này cho thấy SmAPRT cũng biểu hiện ổn định trong cả mô thân và mô quả, vốn không được khảo sát trong nghiên cứu của Gantasala và cộng sự 21 .
Sự biểu hiện phiên mã của gene SmAPRT khi cây cà tím chịu tác động của các yếu tố gây căng thẳng
Giá trị C t của các phản ứng qPCR trong thí nghiệm khảo sát sự biểu hiện phiên mã của gene SmAPRT trong điều kiện cây cà tím chịu tổn thương cơ học hoặc được xử lý với MeJA hay MeSA được thể hiện ở Table 4 . Kết quả phân tích thống kê so sánh giá trị chu kỳ ngưỡng C t giữa nghiệm thức đối chứng và nghiệm thức xử lý bằng phương pháp Student’s t -test (two-tail, unpaired, equal variance) cho thấy không có sự khác biệt có ý nghĩa thống kê giữa mẫu xử lý và mẫu đối chứng (p-value > 0,05) ( Table 7 ). Điều này cho thấy gene SmAPRT biểu hiện ổn định ở tất cả các nghiệm thức của thí nghiệm mô phỏng cây cà tím chịu các tác động từ các yếu tố gây căng thẳng nêu trên.
Năm 2014, nhóm nghiên cứu của Zhou và cộng sự 27 đã khảo sát sự biểu hiện phiên mã của 9 gene giữ nhà ( 18sRNA, CYP, UBQ, GAPDH, TUB, TUA, EF1, ACTIN và RPOB ) trên 4 hệ thống thực nghiệm khác nhau ở cây cà tím S. melongena L. bao gồm các mô khác nhau, xử lý hormone (ABA, MeJA và SA), căng thẳng sinh học (xâm nhiễm tuyến trùng Verticillium dahliae ) và căng thẳng phi sinh học (nóng, lạnh, hạn hán và nhiễm mặn). Dựa trên nghiên cứu này, xét một cách tổng thể, gene mã hóa cho GAPDH và 18S RNA được cho là có biểu hiện ổn định nhất trong các điều kiện thí nghiệm được khảo sát. Tuy nhiên, có thể thấy rõ mức độ ổn định trong sự biểu hiện phiên mã của từng gene giữ nhà thay đổi đáng kể giữa các hệ thống thực nghiệm khác nhau. Nói cách khác, mức biểu hiện phiên mã của các gene có thể ổn định trong điều kiện thí nghiệm này nhưng lại có sự biến động trong những điều kiện khác. Gene mã hóa 18S RNA được đánh giá là có biểu hiện ổn định nhất trong thí nghiệm xử lý hormone, tuy nhiên chỉ được xếp ở vị trí thứ 6 về độ ổn định trong thí nghiệm khảo sát sự biểu hiện theo mô. Ngược lại, gene mã hóa GAPDH đứng đầu về mức độ ổn định trong biểu hiện ở các mô khác nhau của cà tím nhưng được xếp ở vị trí thứ 4 ở thí nghiệm xử lý hormone 27 . Trong báo cáo này, gene SmAPRT không chỉ thể hiện tính ổn định trong biểu hiện ở các mô khác nhau mà còn cả trong điều kiện cây cà tím chịu ảnh hưởng của một số tác động của các tác nhân gây căng thẳng bao gồm xử lý gây tổn thương cơ học, xử lý với hormone (MeJA hoặc MeSA).
KẾT LUẬN
Gene adenine phosphoribosyl transferase (SmAPRT) có mức biểu hiện phiên mã ổn định ở các mô khác nhau của cà tím (p-value = 0,70 > 0,05) hoặc khi cây cà tím ở trong điều kiện căng thẳng như có tổn thương cơ học (p-value = 0,16 > 0,05), hoặc có xử lý với MeJA (p-value = 0,12 > 0,05) hay MeSA (p-value = 0,70 > 0,35) - những hormone liên quan đến con đường truyền tín hiệu căng thẳng. Do vậy, SmAPRT có thể là gene tham chiếu phù hợp cho các phân tích định lượng sự biểu hiện gene bằng phương pháp RT-qPCR trong các điều kiện thí nghiệm nêu trên. Phản ứng qPCR với cặp mồi được thiết kế trong Table 1 đã cho phép nhân bản đặc hiệu một đoạn trình tự DNA có kích thước 103 bp từ bản sao gene SmAPRT với hiệu suất nhân bản (92,7%) trong khoảng khuyến nghị 90‒110%.
PHỤ LỤC
Table 5 , Table 6 , Table 7 , Table 8 , Table 9
LỜI CẢM ƠN
Nhóm nghiên cứu chân thành cảm ơn TS. Trần Kim Cương, Viện Cây ăn quả miền Nam đã cung cấp hạt giống cà tím.
DANH MỤC CÁC TỪ VIẾT TẮT
ABA: abscisic acid
APRT: adenine phosphoribosyl transferase
CYP: cyclophilin
dNTP: deoxynucleotide triphosphate
E%: PCR efficiency
EF1: elongation factor 1-alphaET: ethylene
GAPDH: glyceraldehyde-3-phosphate dehydrogenase
HSD: honestly significant difference
JA: jasmonic acid
MeJA: methyl jasmonate
MeSA: methyl salicylate
RPOB: RNA polymerase beta subunit
RT-qPCR: reverse transcriptase – quantitative polymerase chain reaction
RuBP: ribulose-1,5-bisphosphate carboxylase
SA: salicylic acid
TUA: alpha-tubulin
TUB: beta-tubulin
UBQ: ubiquitin
XUNG ĐỘT LỢI ÍCH
Các tác giả không có xung đột lợi ích nào liên quan đến các kết quả đã công bố.
ĐÓNG GÓP CỦA CÁC TÁC GIẢ
Khuất Lê Uyên Vy lên kế hoạch nghiên cứu, thực hiện các thí nghiệm, thu thập, xử lý dữ liệu và viết bản thảo. Bùi Thị Tường Vi thực hiện các thí nghiệm, thu thập và xử lý dữ liệu. Nguyễn Thị Hồng Thương định hướng và lên kế hoạch nghiên cứu, hướng dẫn nghiên cứu và góp ý bản thảo. Phạm Thị Ánh Hồng đồng hướng dẫn nghiên cứu.
References
- Franklin G, Sheeba C, Lakshmi Sita G. Regeneration of eggplant (Solanum melongena L.) from root explants. In vitro Cellular & Developmental Biology-Plant. 2004;40(2):188-91. . ;:. Google Scholar
- Magioli C, Mansur E. Eggplant (Solanum melongena L.): tissue culture, genetic transformation and use as an alternative model plant. Acta Botanica Brasilica. 2005;19(1):139-48. . ;:. Google Scholar
- Barchi L, Pietrella M, Venturini L, Minio A, Toppino L, Acquadro A, et al. A chromosome-anchored eggplant genome sequence reveals key events in Solanaceae evolution. Scientific Reports. 2019;9(1):1-13. . ;:. PubMed Google Scholar
- Chen L-m, Li X-w, He T-j, Li P-j, Liu Y, Zhou S-x. Comparative biochemical and transcriptome analyses in tomato and eggplant reveal their differential responses to Tuta absoluta infestation. Genomics. 2021;113(4):2108-21. . ;:. PubMed Google Scholar
- Pang Y-Z, Wang Z-H, Guo S-S, Zhang S-S, Zheng L-W, Zhang J-Z. Verticillium dahliae reduces plant growth, constitutively induces antioxidant metabolism and gene expression in eggplant (Solanum melongena L.). Physiological and Molecular Plant Pathology. 2021;114:101641. . ;:. Google Scholar
- Miyatake K, Saito T, Negoro S, Yamaguchi H, Nunome T, Ohyama A. Detailed mapping of a resistance locus against Fusarium wilt in cultivated eggplant (Solanum melongena). Theoretical and Applied Genetics. 2016;129(2):357-67. . ;:. PubMed Google Scholar
- Maffei ME, Arimura G-I, Mithöfer A. Natural elicitors, effectors and modulators of plant responses. Natural Product Reports. 2012;29(11):1288-303. . ;:. PubMed Google Scholar
- Pieterse CM, Van der Does D, Zamioudis C, Leon-Reyes A, Van Wees SC. Hormonal modulation of plant immunity. Annual Review of Cell and Developmental Biology. 2012;28:489-521. . ;:. PubMed Google Scholar
- Howe GA, Jander G. Plant immunity to insect herbivores. Annu Rev Plant Biol. 2008;59:41-66. . ;:. PubMed Google Scholar
- Czechowski T, Stitt M, Altmann T, Udvardi MK, Scheible W-R. Genome-wide identification and testing of superior reference genes for transcript normalization in Arabidopsis. Plant Physiology. 2005;139(1):5-17. . ;:. PubMed Google Scholar
- Bézier A, Lambert B, Baillieul F. Cloning of a grapevine Botrytis‐responsive gene that has homology to the tobacco hypersensitivity‐related hsr203J. Journal of Experimental Botany. 2002;53(378):2279-80. . ;:. PubMed Google Scholar
- Burleigh SH. Relative quantitative RT-PCR to study the expression of plant nutrient transporters in Arbuscular mycorrhizas. Plant Science. 2001;160(5):899-904. . ;:. Google Scholar
- Foss DL, Baarsch MJ, Murtaugh MP. Regulation of hypoxanthine phosphoribosyltransferase, glyceraldehyde‐3‐phosphate dehydrogenase and β‐actin mRNA expression in porcine immune cells and tissues. Animal Biotechnology. 1998;9(1):67-78. . ;:. PubMed Google Scholar
- Schmittgen TD, Zakrajsek BA. Effect of experimental treatment on housekeeping gene expression: validation by real-time, quantitative RT-PCR. Journal of Biochemical and Biophysical Methods. 2000;46(1-2):69-81. . ;:. Google Scholar
- Thellin O, Zorzi W, Lakaye B, De Borman B, Coumans B, Hennen G. Housekeeping genes as internal standards: use and limits. Journal of Biotechnology. 1999;75(2-3):291-5. . ;:. Google Scholar
- Suzuki T, Higgins PJ, Crawford DR. Control selection for RNA quantitation. Biotechniques. 2000;29(2):332-7. . ;:. PubMed Google Scholar
- Lee PD, Sladek R, Greenwood CM, Hudson TJ. Control genes and variability: absence of ubiquitous reference transcripts in diverse mammalian expression studies. Genome Research. 2002;12(2):292-7. . ;:. PubMed Google Scholar
- Remans T, Keunen E, Bex GJ, Smeets K, Vangronsveld J, Cuypers A. Reliable gene expression analysis by reverse transcription-quantitative PCR: reporting and minimizing the uncertainty in data accuracy. The Plant Cell. 2014;26(10):3829-37. . ;:. PubMed Google Scholar
- Chapman JR, Waldenström J. With reference to reference genes: a systematic review of endogenous controls in gene expression studies. PloS one. 2015;10(11):e0141853. . ;:. PubMed Google Scholar
- Owczarzy R, Tataurov AV, Wu Y, Manthey JA, McQuisten KA, Almabrazi HG. IDT SciTools: a suite for analysis and design of nucleic acid oligomers. Nucleic Acids Research. 2008;36(suppl-2):W163-W9. . ;:. PubMed Google Scholar
- Gantasala NP, Papolu PK, Thakur PK, Kamaraju D, Sreevathsa R, Rao U. Selection and validation of reference genes for quantitative gene expression studies by real-time PCR in eggplant (Solanum melongena L). BMC Research Notes. 2013;6(1):1-11. . ;:. PubMed Google Scholar
- Falara V, Alba JM, Kant MR, Schuurink RC, Pichersky E. Geranyllinalool synthases in solanaceae and other angiosperms constitute an ancient branch of diterpene synthases involved in the synthesis of defensive compounds. Plant Physiology. 2014;166(1):428-41. . ;:. PubMed Google Scholar
- Ament K, Van Schie CC, Bouwmeester HJ, Haring MA, Schuurink RC. Induction of a leaf specific geranylgeranyl pyrophosphate synthase and emission of (E,E)-4,8,12-trimethyltrideca-1,3,7,11-tetraene in tomato are dependent on both jasmonic acid and salicylic acid signaling pathways. Planta. 2006;224(5):1197-208. . ;:. PubMed Google Scholar
- Svec D, Tichopad A, Novosadova V, Pfaffl MW, Kubista M. How good is a PCR efficiency estimate: Recommendations for precise and robust qPCR efficiency assessments. Biomolecular Detection and Quantification. 2015;3:9-16. . ;:. PubMed Google Scholar
- Töwe S, Kleineidam K, Schloter M. Differences in amplification efficiency of standard curves in quantitative real-time PCR assays and consequences for gene quantification in environmental samples. Journal of Microbiological Methods. 2010;82(3):338-41. . ;:. PubMed Google Scholar
- Rogers‑Broadway KR, Karteris E. Amplification efficiency and thermal stability of qPCR instrumentation: Current landscape and future perspectives. Experimental and Therapeutic Medicine. 2015;10(4):1261-4. . ;:. PubMed Google Scholar
- Zhou X, Liu J, Zhuang Y. Selection of appropriate reference genes in eggplant for quantitative gene expression studies under different experimental conditions. Scientia Horticulturae. 2014;176:200-7. . ;:. Google Scholar





