

 Open Access
Open Access Abstract
In the emergency that the Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) has globally spread, vaccine is one of the most indispensable defense lines to repulse the resulting Coronavirus Disease 2019 (COVID-19) pandemic. The effective immunization frequently requires appropriate adjuvants which play a key role to activate the dendritic cell maturation. Indeed, the antigen presentation of dendritic cells indisputably needs the collaboration of co-stimulatory molecules which are only expressed by mature dendritic cells to trigger T cell response. In this review, we collectively summarized well-known adjuvants used for vaccination as well as their characteristics and immunostimulatory effects on both of humoral and cell-mediated immune response. Besides, the applicability of adjuvants in COVID-19 vaccination and accompanying challenges were also discussed. Generally, despite of certain successes of available licensed adjuvants in the vaccination, new generations of novel adjuvants would absolutely be required to produce effective vaccines for other inevitable pathogenic organisms in the future.
MỘT SỐ VACCINE PHÒNG COVID-19 NỔI BẬT
Đứng trước thực trạng virus gây bệnh viêm phổi cấp SARS-CoV-2 đang lây lan với tốc độ nhanh chóng trên toàn cầu, vaccine là một trong những tuyến phòng thủ quan trọng mà con người có thể sử dụng để kìm hãm sự lây lan của virus, cũng như giúp người được tiêm vaccine tránh được những triệu chứng nặng và trường hợp tử vong nếu nhiễm phải virus. Hiện nay, nhiều loại vaccine phòng COVID-19 đã ra đời dựa trên nhiều công nghệ khác nhau, bao gồm: vaccine virus bất hoạt (Vero Cell của Sinopharm hay CoronaVac của Sinovac Biotech) 1 , 2 , vaccine vector virus (Vaxzevria của Oxford/AstraZeneca hay Sputnik V của Viện Nghiên cứu Dịch tễ học và Vi sinh vật học Gamaleya, Nga) 3 , vaccine mRNA (Comirnaty của Pfizer/BioNTech hay Spikevax của Moderna) 3 , hay vaccine tiểu phần (NVX-CoV2373 của Novavax hay Nanocovax của Nanogen đã hoàn thành đánh giá thử nghiệm lâm sàng giai đoạn 3a) 4 , 5 . Nhìn chung, các vaccine có chung cách tiếp cận là sử dụng những kháng nguyên từ SARS-CoV-2 để gây đáp ứng trong cơ thể nhằm tạo kháng thể kháng lại virus. Trong đó, kháng nguyên đóng vai trò chính trong quá trình gây đáp ứng là protein gai S (spike protein) trên hạt virus giữ nhiệm vụ tương tác với thụ thể angiotensin-converting enzyme 2 (ACE2) trên nguyên bào sợi phổi 3 , 6 . Do đó, việc gây đáp ứng để hệ miễn dịch tạo kháng thể trung hòa protein gai S giúp khóa tương tác giữa virus và thụ thể trên tế bào chủ, từ đó ngăn virus xâm nhập.
VAI TRÒ CỦA TÍN HIỆU TỪ CÁC KIỂU MẪU PHÂN TỬ CỦA VI SINH VẬT TRONG ĐÁP ỨNG MIỄN DỊCH THÍCH ỨNG
Trong các vaccine tiểu phần, mặc dù kháng nguyên (vai trò cung cấp tín hiệu thứ 1 trong quá trình đáp ứng miễn dịch) là thành phần chính của vaccine, tuy nhiên hiệu quả gây đáp ứng sẽ không cao nếu kháng nguyên không được đi kèm với tá dược miễn dịch... Trong quá trình đáp ứng miễn dịch, các tế bào tua (dendritic cell/DC) trình diện các đoạn peptide từ kháng nguyên đã được thực bào trên các phức hợp tương hợp mô chính (major histocompatibility complex/MHC) trên bề mặt để tế bào T nhận diện thông qua thụ thể tế bào T (T-cell receptor/TCR) 7 . Chỉ khi được hoạt hóa bằng quá trình trình diện kháng nguyên từ tế bào tua, tế bào T mới có thể tiếp tục tham gia vào các bước tiếp theo trong quá trình đáp ứng của hệ miễn dịch thích ứng (adaptive immune response) 7 . Tuy nhiên, quá trình hoạt hóa tế bào T đòi hỏi tín hiệu của các phân tử đồng kích thích (tín hiệu thứ 2) là protein B7 (gồm hai dạng là B7-1 và B7-2 tương tác với đồng thụ thể kích thích CD28 trên tế bào T để truyền tín hiệu hoạt hóa) trên tế bào tua 7 , 8 , 9 ( Figure 1 A). Nếu quá trình trình diện kháng nguyên thiếu sự hỗ trợ của phân tử đồng kích thích, hiệu quả hoạt hóa tế bào T rất thấp ( Figure 1 B), dẫn đến quá trình đáp ứng miễn dịch thích ứng kém hiệu quả 7 , 10 . Đến nay, các nghiên cứu đã làm sáng tỏ nguồn gốc tín hiệu thứ 2 của các phân tử đồng kích thích xuất phát từ quá trình tế bào tua nhận diện các kiểu mẫu phân tử liên quan tới vi sinh vật xâm nhiễm (pathogen-associated molecular pattern/PAMP) 7 . PAMP là các dạng phân tử xuất hiện chung ở số lượng lớn những loài vi sinh vật, chẳng hạn lipopolysaccharide (LPS) ở vi khuẩn gram âm, RNA mạch đôi (dsRNA) ở một số virus, peptidoglycan ở vi khuẩn, glucan ở nấm. Khi vi sinh vật xâm nhập vào cơ thể, các PAMP của chúng được nhận diện bởi các thụ thể nhận diện kiểu mẫu phân tử (pattern recognition receptor/PRR) của các tế bào của hệ miễn dịch bẩm sinh (innate immune system), trong đó có tế bào tua 7 , 11 , 12 . Các PRR có thể là những thụ thể đính màng như các Toll-like receptor (TLR) 13 , các C-type lectin receptor (CLR) 14 hay những thụ thể nội bào như retinoic acid-inducible gene I (RIG-I), melanoma differentiation-associated protein 5 (MDA5) 15 , 16 . Quá trình nhận diện các PAMP thông qua PRR kích hoạt những con đường tín hiệu nội bào hoạt hóa các nhân tố phiên mã (như nuclear factor κB/NF-κB, interferon regulatory factor/IRF 3, c-Jun N-terminal kinase 1/JNK1) 17 , 18 , 19 để thúc đẩy tế bào tua trưởng thành, biểu hiện các phân tử đồng kích thích, và tiết các cytokine như interleukin (IL)-2, IL-12 kích hoạt hệ miễn dịch thích ứng 20 . Một số cặp PAMP/PRR điển hình bao gồm: lipoteichoic acid/TLR2, peptidoglycan/TLR1-TLR2, LPS/TLR4, flagellin/TLR5, RNA mạch đôi (double-stranded RNA/dsRNA)/TLR3, RNA virus ngắn/RIG-I, RNA virus dài/MDA5, β-glucan/dectin 1, α-mannan/dectin 2 ( Figure 2 ) 16 . Do đó, trong các loại vaccine, tá dược miễn dịch được sử dụng thường là các dạng PAMP thu nhận từ vi sinh vật (trừ một số trường hợp ngoại lệ) giúp thúc đẩy quá trình trưởng thành của tế bào tua để cung cấp tín hiệu thứ 2 cho quá trình đáp ứng miễn dịch thích ứng.
Figure 1 . Quá trình trình diện kháng nguyên của tế bào tua khi có (A) và không có (B) phân tử đồng kích thích

Figure 2 . Quá trình nhận diện các PAMP bằng TLR (A), PRR nội bào (B), và CLR (C) 16 .

CÁC TÁ DƯỢC MIỄN DỊCH HỖ TRỢ ĐÁP ỨNG MIỄN DỊCH DỊCH THỂ (HUMORAL IMMUNE RESPONSE)
Đáp ứng miễn dịch dịch thể cùng với đáp ứng miễn dịch qua trung gian tế bào là hai dạng đáp ứng đặc trưng của hệ miễn dịch thích ứng. Quá trình đáp ứng miễn dịch dịch thể bắt đầu khi tế bào T CD4 + , còn gọi là tế bào T giúp đỡ (helper T cell/T H ) gồm các dạng chính là T H 1, T H 2, T H 17, được hoạt hóa sau khi nhận diện kháng nguyên được trình diện trên phân tử MHC lớp II (MHC-II) của tế bào tua 7 . Sau đó, các tế bào T giúp đỡ tiếp tục hoạt hóa tế bào B mang thụ thể đặc hiệu kháng nguyên nhân dòng và biệt hóa thành các tương bào tiết kháng thể nhận diện và trung hòa các vi sinh vật xâm nhiễm 7 . Đến nay, một số các tá dược miễn dịch hỗ trợ quá trình đáp ứng miễn dịch dịch thể đã được nghiên cứu và phát triển.
Tá dược miễn dịch nhôm (Alum)
Tá dược miễn dịch nhôm (các muối không tan của nhôm, phổ biến nhất là aluminium hydroxide, aluminium phosphate, aluminium oxyhydroxide) là một tá dược miễn dịch đặc biệt vì không có bản chất là PAMP từ vi sinh vật như những tá dược miễn dịch khác 21 , 22 , 23 . Một số vaccine COVID-19 sử dụng tá dược miễn dịch nhôm là Nanocovax (Nanogen, Việt Nam) hay MVC-COV1901 (Medigen, Đài Loan) 24 . Mặc dù cách thức kích thích tế bào T CD4 + của tá dược miễn dịch nhôm vẫn chưa được hiểu hết, tuy nhiên một số cơ chế đã được đề xuất. Cụ thể, ion Al 3+ trong tinh thể muối nhôm có khả năng tương tác với những nhóm chức tích điện âm trên protein kháng nguyên, từ đó dẫn đến quá trình trao đổi giữa kháng nguyên với những anion trong tinh thể muối nhôm làm thúc đẩy kháng nguyên hấp phụ lên bề mặt tinh thể ( Figure 3 ). Từ đó, kháng nguyên khi đưa vào cơ thể được giải phóng chậm từ tinh thể muối nhôm giúp quá trình đáp ứng miễn dịch được kéo dài liên tục theo thời gian 25 . Cơ chế thứ hai được các nghiên cứu đề cập là khả năng gây tổn thương mô của nhôm. Trong đó, các tế bào chết ở mô bị tổn thương tại vị trí gây đáp ứng giải phóng ra các kiểu mẫu phân tử liên quan đến tổn thương (damage-associated molecular pattern/DAMP) như adenosine triphosphate (ATP), uric acid, DNA, và các thành phần này hoạt hóa hệ miễn dịch thích ứng 26 , 27 , 28 , 29 . Trong đó, ATP và uric acid cảm ứng con đường tín hiệu viêm của các phân tử NACHT, LRR and PYD domains-containing protein 3 (NLRP3), còn các DNA giải phóng từ tế bào chết sau khi bị tế bào tua thực bào được nhận diện bởi enzyme nội bào cyclic guanosine monophosphate (GMP)-adenosine monophosphate (AMP) synthase (cGAS) để cảm ứng stimulator of interferon gene (STING), là protein thiết yếu của tính miễn dịch bẩm sinh. NLRP3 và STING sau khi được cảm ứng sẽ hoạt hóa các nhân tố phiên mã như NF-κB và IRF3 giúp tăng cường biểu hiện các cytokine giữ vai trò kích thích hệ miễn dịch thích ứng 16 , 26 , 30 , 31 . Tuy nhiên, nhược điểm lớn nhất của tá dược miễn dịch nhôm là khả năng kích thích tế bào B chuyển lớp kháng thể sang immunoglobulin (Ig) E 26 (dạng kháng thể giữ vai trò ngăn chặn kí sinh trùng và liên quan đến dị ứng). Không như kháng thể IgG, IgE rất ít phân bố trong máu mà thường chỉ có nhiều ở màng nhầy, do đó khả năng nhận diện và trung hòa vi sinh vật trong máu của IgE rất thấp 32 . Cơ chế tá dược miễn dịch nhôm giúp tế bào B chuyển lớp kháng thể sang IgE đã được chứng minh là bởi IL-6 (một trong số các cytokine được tiết bởi tế bào tua nhờ con đường NF-κB và IRF3) sẽ kích thích tế bào T giúp đỡ biệt hóa thành T H 2 33 , 34 , 35 . Cuối cùng, các tế bào T H 2 tiết các cytokine như IL-4 kích thích tế bào B chuyển lớp kháng thể và tiết IgE 26 , 36 , 37 , 38 , 39 . Ngoài ra, tá dược miễn dịch nhôm cũng có nhược điểm phụ là khả năng gây độc cho tế bào và mô 40 .

Tá dược miễn dịch nhũ hóa (emulsion adjuvant)
Giống như nhôm, tá dược miễn dịch nhũ hóa cũng không phải là PAMP từ vi sinh vật. Trong đó, squalene (một dạng dầu với bản chất là hydrocarbon không no, có tính tương thích sinh học và phân hủy sinh học và thường được sử dụng như một dạng dưỡng chất bổ sung cho cơ thể) là chất nhũ hóa phổ biến được sử dụng làm tá dược miễn dịch. Với tính kỵ nước cao, squalene được sử dụng làm chất nhũ hóa để tạo nên những cấu trúc hạt có khả năng mang và giải phóng chậm kháng nguyên tại vị trí gây đáp ứng giúp quá trình đáp ứng miễn dịch được diễn ra liên tục trong thời gian dài, từ đó nâng cao hiệu quả tiết kháng thể của tế bào B 41 , 42 . Ngoài ra, trong vaccine, squalene còn được nhũ tương hóa bằng một số chất hoạt động bề mặt như Tween 80 hay Span 85 (tá dược miễn dịch MF59) để giúp những hạt squalene tan trong nước tốt hơn 43 ( Figure 4 ). Cơ chế hỗ trợ đáp ứng miễn dịch thích ứng của MF59 cũng đã được chứng minh 16 . Cụ thể, MF59 kích thích đại thực bào, bạch cầu đơn nhân, bạch cầu hạt tiết các chemokine như C-C motif chemokine ligand (CCL) 2, CCL4, CCL5, C-X-C motif chemokine ligand (CXCL) 8, từ đó thu hút các tế bào miễn dịch đến vị trí gây đáp ứng và tiếp tục tiết các chemokine này. Quá trình khuếch đại tín hiệu chemokine kích thích tế bào tua cùng một số tế bào miễn dịch khác (thông qua tương tác giữa chemokine với các thụ thể tương ứng trên tế bào) di chuyển về các hạch bạch huyết để trình diện kháng nguyên cho tế bào T 44 , 45 . Ngoài ra, MF59 cũng được chứng minh có khả năng gây tổn thương mô để giải phóng những thành phần trong tế bào như ATP, uric acid giúp kích thích đáp ứng miễn dịch thích ứng tương tự cơ chế của tá dược miễn dịch nhôm 46 , 47 . Dạng tá dược miễn dịch nhũ hóa cải tiến khác là Adjuvant System 03 (AS03), một sự kết hợp giữa MF59 và α-tocopherol, một dạng của vitamin E ( Figure 4 ). Trong các thử nghiệm trên chuột, α-tocopherol thúc đẩy các tế bào miễn dịch tiết CCL2, CCL3, CXCL1, IL-6, granulocyte colony-stimulating factor (G-CSF) để kích thích các bạch cầu thực bào kháng nguyên và di chuyển về hạch bạch huyết để hỗ trợ đáp ứng miễn dịch thích ứng 48 . Trong một nghiên cứu thử nghiệm vaccine H5N1 trên người, tá dược miễn dịch AS03 cho thấy khả năng kích thích sự biểu hiện của CXCL10 và interferon (IFN)-γ giúp tế bào tua, bạch cầu trung tính, bạch cầu đơn nhân tăng biểu hiện của các gen mã hóa MHC, các enzyme phân cắt peptide như endoplasmic reticulum aminopeptidase 2 (ERAP2), tripeptidyl-peptidase 2 (TPP2) 49 cần thiết cho quá trình xử lý và trình diện kháng nguyên.

Tá dược miễn dịch LPS
Tá dược miễn dịch LPS là một trong những tá dược miễn dịch điển hình có nguồn gốc là PAMP của vi sinh vật. LPS, còn gọi là nội độc tố (endotoxin), là những phân tử lipid liên kết các chuỗi polysaccharide có nguồn gốc từ màng ngoài (outer membrane) của vi khuẩn gram âm. LPS kích thích tế bào tua trưởng thành và biểu hiện phân tử đồng kích thích B7-1 bằng con đường tín hiệu của TLR4 50 , 51 , từ đó hoạt hóa hệ miễn dịch thích ứng đáp ứng theo hướng T H 1 giúp kích hoạt tế bào B tiết kháng thể IgG 52 . Tuy có khả năng kích thích mạnh hệ miễn dịch thích ứng, LPS vẫn mang độc tính vì có bản chất là nội độc tố của vi khuẩn, trong đó nghiêm trọng nhất là khả năng gây sốc nhiễm trùng và tử vong 53 . Do đó, để ứng dụng làm tá dược miễn dịch trong vaccine, LPS thường được biến đổi để loại bỏ độc tính. Dạng LPS không độc được sử dụng nhiều nhất để làm tá dược miễn dịch là monophosphoryl lipid A (MPL) được tạo ra bằng phương pháp thủy phân trong acid các phân tử lipid A từ vi khuẩn để loại bớt một nhóm phosphate ở vị trí 1′ của gốc đường glucosamine và loại các chuỗi bên polysaccharide, hoặc thậm chí có thể loại bớt một chuỗi acid béo ở vị trí 3′ trên glucosamine tạo ra sản phẩm 3- O -desacyl-4′-monophosphoryl lipid A, còn gọi là 3-deacylphosphorylated hexaacyl disaccharide (3D-PHAD) ( Figure 5 A) 54 , 55 , 56 . Các nghiên cứu cho thấy MPL có độc tính giảm hơn 1000 lần so với các dạng LPS gốc 54 do mất đi một nhóm phosphate nên làm giảm sự biểu hiện của các cytokine tiền viêm liên quan đến sốc nhiễm trùng 57 . Tuy nhiên, dù độc tính giảm rất nhiều, nhưng MPL vẫn giữ được khả năng kích thích đáp ứng miễn dịch thích ứng 58 , 59 . Trong một số nghiên cứu, 3D-PHAD được sử dụng ở dạng hấp phụ trên tá dược miễn dịch nhôm thông qua nhóm phosphate tích điện âm (tạo thành hỗn hợp tá dược miễn dịch AS04) để nâng cao hiệu quả hỗ trợ đáp ứng miễn dịch thích ứng ( Figure 5 B) 60 , 61 . Tương tự với AS04, AS01 cũng là một tá dược miễn dịch sử dụng 3D-PHAD nhưng được kết hợp với saponin QS-21 trong cấu trúc hạt liposome 62 , 63 ( Figure 5 B). QS-21 đã được chứng minh là có khả năng kích thích các tế bào tua nuôi cấy in vitro (được biệt hóa từ tế bào THP-1) trưởng thành thông qua sự tăng biểu hiện của IL-6, IL-8, tumor necrosis factor (TNF), MHC-II, và phân tử đồng kích thích B7-2. Tác động này đã được chứng minh là thông qua hai cơ chế: một là sau khi các hạt liposome QS-21 được nhập bào (endocytosis), với bản chất là saponin, các QS-21 có khả năng làm tăng tính thấm của các túi lysosome làm giải phóng cathepsin B giúp hoạt hóa NF-κB; hai là QS-21 có khả năng hoạt hóa con đường tín hiệu của spleen tyrosine kinase (Syk) 64 .
Figure 5 . Phản ứng tổng hợp 3D-PHAD từ LPS (A) và cấu trúc các tá dược miễn dịch AS01 và AS04 (B)

Tá dược miễn dịch cytosine-phosphate-guanine (CpG)
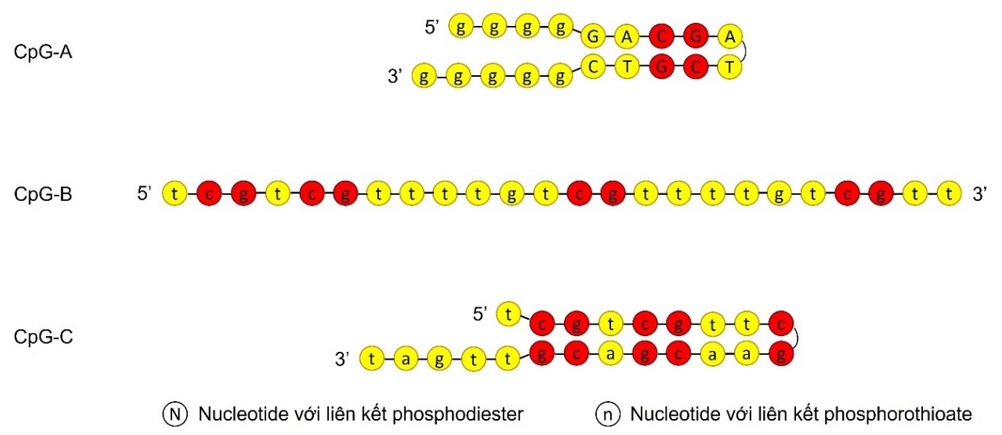
Các oligodeoxynucleotide (ODN) CpG ở vi sinh vật có khả năng kích thích mạnh hệ miễn dịch thích ứng đáp ứng theo hướng T H 1. Các đoạn CpG này sau khi được tế bào tua thực bào tương tác với TLR9 đính màng endosome để kích hoạt con đường tín hiệu phụ thuộc myeloid differentiation factor 88 (MyD88) giúp hoạt hóa các nhân tố phiên mã như IRF3, IRF7 thúc đẩy tế bào tua trưởng thành và tiết các cytokine như IFN-α và IL-6 65 . Hiện nay, các tá dược miễn dịch CpG được sử dụng trong nghiên cứu vaccine đều là những đoạn ODN mạch đơn được tổng hợp hóa học mang biến đổi phosphorothioate để kháng lại tác động phân cắt của các nuclease nội bào và được chia làm ba loại là CpG-A, CpG-B, và CpG-C với trình tự và cấu trúc riêng ( Figure 6 ) 65 , 66 , 67 , 68 . Trong đó, CpG-A thường chỉ mang một hoặc hai dinucleotide CG ở đoạn trình tự bắt cặp đối ngẫu (palindromic motif) và có thêm đuôi poly-G mang biến đổi phosphorothioate ở đầu 3′ và 5′ 65 , 66 , 68 . Khác với CpG-A, CpG-B không có đuôi poly-G, thường tồn tại ở dạng mạch thẳng tuy một số trường hợp có thể ở dạng tự bắt cặp, mang biến đổi phosphorothioate trên toàn bộ trình tự và có nhiều CG hơn 65 , 66 , 67 , 68 . CpG-C là trình tự mang những đặc điểm của cả hai dạng CpG-A và CpG-B với trình tự bắt cặp đối ngẫu chứa nhiều CG, mang biến đổi phosphorothioate trên toàn bộ trình tự và không có đuôi polyG 65 , 66 , 67 , 68 . Cách thức kích thích đáp ứng miễn dịch của CpG phụ thuộc vào việc CpG tồn tại ở dạng monomer hay multimer (được quyết định bởi trình tự và cấu trúc của CpG) 69 . Trong đó, CpG-B (tồn tại ở dạng monomer) sau khi bị thực bào được chuyển vào các túi endosome muộn biểu hiện lysosome-associated membrane protein-1 (LAMP-1), sau đó hoạt hóa tế bào tua trưởng thành (tăng mức biểu hiện B7-1 và B7-2) nhưng lại không tiết IFN-α 69 . Ngược lại, CpG-A, với khả năng kết cụm thành dạng multimer nhờ có đuôi poly-G, sẽ được chuyển vào các túi endosome biểu hiện transferrin receptor (TfR), từ đó kích thích tế bào tua tiết IFN-α nhưng lại không trưởng thành (mức biểu hiện các phân tử B7 rất thấp) 69 . CpG-C, tồn tại ở dạng dimer do mang đặc điểm lai của CpG-A và CpG-B, có khả năng kích thích tế bào tua trưởng thành lẫn tiết IFN-α vì có thể được chuyển vào cả hai dạng túi endosome trên 69 . Do đó, cách thức tá dược miễn dịch CpG hoạt hóa tế bào tua phụ thuộc vào dạng túi endosome diễn ra tương tác giữa CpG và TLR9. Tuy nhiên, cơ chế phân tử của vấn đề này vẫn chưa được xác định. Hiện nay, tá dược miễn dịch CpG đã được sử dụng rất nhiều trong nghiên cứu vaccine, trong đó có cả vaccine cho COVID-19 70 , 16 . Tuy nhiên, mặc dù có rất nhiều loại CpG đã được nghiên cứu, chỉ có CpG 1018, một dạng CpG-B dài 22 nucleotide với trình tự 5′-TGACTGTGAACGTTCGAGATGA-3′ 66 , là tá dược miễn dịch CpG duy nhất được cấp phép sử dụng cho vaccine trên người 70 , 16 .
Tá dược miễn dịch flagellin
Flagellin là một protein hình cầu có khả năng tự liên kết để tạo thành cấu trúc sợi tiên mao (lông roi) ở một số vi khuẩn gram âm, thí dụ như Escherichia coli hay Salmonella 71 . Flagellin được chứng minh là phối tử của TLR5 trên các tế bào trình diện kháng nguyên. Hiện nay, flagellin bắt đầu được quan tâm nghiên cứu để có thể phát triển thành một dạng tá dược miễn dịch mới cho vaccine. Flagellin có đặc điểm nổi bật là khả năng kích thích mạnh hệ miễn dịch thích ứng đáp ứng theo hướng T H 1 nhưng lại ít gây ra tác dụng phụ, thí dụ khả năng gây độc, như các tá dược miễn dịch nhôm hay LPS 72 . Cơ chế hoạt hóa tế bào tua của flagellin cũng đã được hiểu rõ. Flagellin có thể tương tác với thụ thể TLR5 dạng homodimer để hoạt hóa NF-κB theo con đường tín hiệu phụ thuộc MyD88, hoặc tương tác với thụ thể heterodimer TLR5/TLR4 để hoạt hóa IRF3 theo con đường tín hiệu phụ thuộc TIR-domain-containing adapter-inducing interferon-β (TRIF) 73 , 74 , 75 . Cả hai nhân tố phiên mã NF-κB và IRF3 sau đó đều cảm ứng sự biểu hiệu của các cytokine tiền viêm quan trọng trong đáp ứng miễn dịch thích ứng như IL-1β, IL-12, INF-β 72 . Tín hiệu của TLR5 kích hoạt bởi flagellin cũng giúp tế bào tua trưởng thành, tăng cường biểu hiện MHC-II và các phân tử đồng kích thích B7, đồng thời kích thích tế bào tua tiết các chemokine như CCL2, CCL3, CCL4, CXCL1, CXCL2, CXCL8 hướng hóa tế bào tua, đại thực bào, bạch cầu di chuyển tới hạch bạch huyết để hỗ trợ đáp ứng miễn dịch thích ứng 76 . Những nghiên cứu trên cho thấy flagellin là một trong những ứng viên tá dược miễn dịch tiềm năng cho đáp ứng T H 1 77 . Tuy nhiên, hạn chế lớn nhất của flagellin là tính sinh miễn dịch vì đây là protein của vi khuẩn, do đó có thể cạnh tranh đáp ứng của hệ miễn dịch với các kháng nguyên của vaccine. Từ đó, một số nghiên cứu đã tiến hành tạo các phiên bản thu nhỏ của flagellin dưới dạng protein tái tổ hợp để giảm tính sinh miễn dịch nhưng vẫn giữ được khả năng tương tác với TLR5 của flagellin nhằm ứng dụng làm tá dược miễn dịch trong vaccine 78 .
TÁ DƯỢC MIỄN DỊCH HỖ TRỢ ĐÁP ỨNG MIỄN DỊCH QUA TRUNG GIAN TẾ BÀO (CELL-MEDIATED IMMUNE RESPONSE)
Khác với đáp ứng miễn dịch dịch thể cần sự giúp đỡ của tế bào T CD4 + , tế bào T giữ vai trò chính trong đáp ứng miễn dịch qua trung gian tế bào là T CD8 + (còn gọi là tế bào T gây độc). Trong đó, các tế bào nhiễm trình diện các peptide từ kháng nguyên của vi sinh vật đã xâm nhập tế bào trên các phân tử MHC lớp I (MHC-I). Sau khi nhận diện kháng nguyên lạ được trình diện trên MHC-I bằng TCR, tế bào T CD8 + tiết các protein gây độc để tiêu diệt tế bào nhiễm 7 . Trong một số bệnh truyền nhiễm gây ra bởi vi sinh vật tăng trưởng nội bào, đáp ứng miễn dịch dịch thể tỏ ra kém hiệu quả do kháng thể không thể tiếp cận được vi sinh vật ẩn náu bên trong tế bào. Khi đó, đáp ứng miễn dịch qua trung gian tế bào bởi tế bào T CD8 + sẽ phát huy hiệu quả để tiêu diệt tế bào nhiễm giúp ngăn vi sinh vật phát triển. Quá trình hoạt hóa tế bào T CD8 + cũng cần tế bào tua hỗ trợ nhưng kháng nguyên sẽ được trình diện trên MHC-I của tế bào tua 7 . Tuy nhiên, trong tế bào tua, chỉ có các kháng nguyên nội bào mới được trình diện trên MHC-I, còn các kháng nguyên được thực bào trong các túi phagosome đều được trình diện theo con đường MHC-II để kích hoạt miễn dịch dịch thể. Do đó, muốn kích thích miễn dịch qua trung gian tế bào, vaccine cần có chiến lược phù hợp để kích thích tế bào tua trình diện chéo kháng nguyên (antigen cross-presentation) từ MHC-II sang MHC-I 7 , 79 , 80 , 81 . Đây thực sự là một thách thức lớn khi hiện nay chỉ có rất ít vaccine làm được điều này 16 . Một trong số hiếm hoi các vaccine gây được đáp ứng miễn dịch qua trung gian tế bào là vaccine yellow fever 17D (YF-17D) ngừa bệnh sốt vàng da 82 .
Tá dược miễn dịch dsRNA
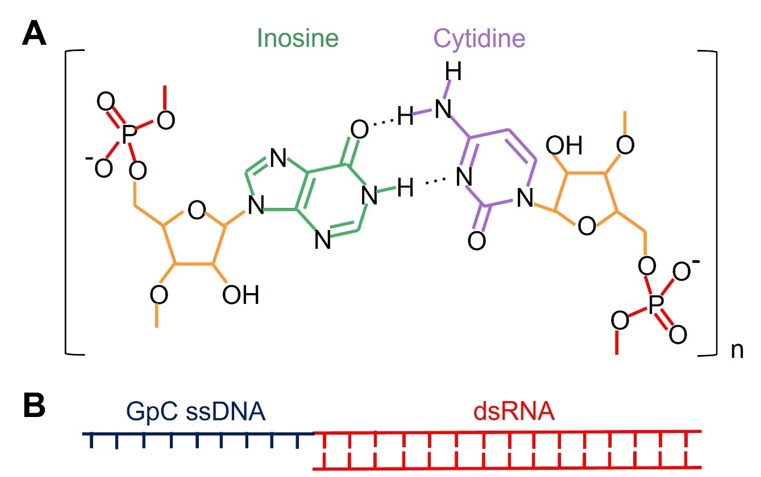
Các phân tử dsRNA là kiểu mẫu đặc trưng ở một số virus có khả năng nổi bật là kích hoạt được đáp ứng của cả tế bào T H 1 lẫn tế bào T CD8 + . Hai dạng dsRNA được sử dụng phổ biến nhất để làm tá dược miễn dịch trong nghiên cứu vaccine là polyinosinic-polycytidylic acid, gọi tắt là poly(I:C) ( Figure 7 A) 83 , 84 , 85 , và guanine-phosphate-cytosine (GpC) DNA-capped dsRNA, còn có tên gọi khác là ARNAX ( Figure 7 B) 86 , 87 , 88 . Trong đó, poly(I:C) là dạng dsRNA tổng hợp hóa học sử dụng hai loại ribonucleotide là inosine và cytidine (không thuộc bốn loại ribonucleotide của RNA trong tự nhiên) có khả năng bắt cặp bổ sung với nhau, còn ARNAX là các đoạn dsRNA được biến đổi để gắn thêm một đoạn DNA GpC mạch đơn ở một đầu 5′. Cơ chế hoạt hóa tế bào T H 1 của các kiểu mẫu dsRNA đã được hiểu rõ là nhờ khả năng kích thích tế bào tua trưởng thành thông qua tương tác với thụ thể TLR3 để hoạt hóa NF-κB và IRF3 thông qua con đường tín hiệu phụ thuộc TRIF 88 . Các nghiên cứu cũng đã chứng minh poly(I:C) có khả năng hoạt hóa tế bào tua trưởng thành với sự tăng biểu hiện của các phân tử B7, MHC-II, và IFN loại I (type I IFN) để hoạt hóa tế bào T CD4 + 89 , 90 . Ngược lại, cơ chế hoạt hóa tế bào T CD8 + của dsRNA vẫn chưa được hiểu rõ. Tuy nhiên, một số nghiên cứu cho thấy cách thức dsRNA thúc đẩy tế bào tua trình diện chéo kháng nguyên để hoạt hóa tế bào T CD8 + cũng phụ thuộc vào con đường tín hiệu TLR3-IRF3 khi chứng minh tá dược miễn dịch ARNAX mất khả năng kích thích đáp ứng của tế bào T CD8 + ở chuột bị loại gene Tlr3 , Ticam1 , Irf3 , trong khi đó khả năng hoạt hóa tế bào T CD8 + của ARNAX vẫn được giữ nguyên khi loại những gene không thuộc con đường tín hiệu của TLR3 như Mavs hay Myd88 91 . Mặc dù cơ chế chưa được hiểu hết, nhiều nghiên cứu vẫn cho thấy dsRNA có khả năng kích thích mạnh tế bào T CD8 + đáp ứng và hình thành tế bào nhớ 92 , thậm chí ngay cả khi không có sự giúp đỡ của tế bào T CD4 + 93 .
Tá dược miễn dịch imidazoquinoline (IMQ)
Các hợp chất IMQ đã được chứng minh có đặc tính của một tá dược miễn dịch thông qua khả năng tương tác với TLR7 và TLR8 là hai dạng thụ thể nhận diện kiểu mẫu ssRNA (thuộc nhóm thụ thể đính màng endosome cùng với TLR3 và TLR9) ở tế bào trình diện kháng nguyên mặc dù IMQ không phải RNA 94 , 95 . Các phân tích tin sinh học đã chứng minh mô hình tương tác giữa một số IMQ và TLR7 95 . Cơ chế TLR7/TLR8 hoạt hóa tế bào tua cũng đã được hiểu rõ là thông qua con đường tín hiệu của MyD88 để hoạt hóa các nhân tố phiên mã NF-κB và IRF7 96 , 97 , 98 . Giống với TLR3, tín hiệu từ TLR7/TLR8 có thể hoạt hóa đáp ứng của cả miễn dịch dịch thể lẫn miễn dịch qua trung gian tế bào 99 . Thí dụ, trong nghiên cứu của Wille-Reece và cộng sự 100 , resiquimod (R-848), một dạng hợp chất IMQ, tỏ ra kém hiệu quả trong việc hoạt hóa đáp ứng của tế bào T H 1 và T CD8 + ở chuột với kháng nguyên protein Gag của human immunodeficiency virus 1 (HIV-1) khi ở dạng tá dược miễn dịch tự do. Tuy nhiên, khi gắn kết R-848 với Gag, hiệu quả đáp ứng của tế bào T H 1 và T CD8 + đặc hiệu kháng nguyên đã được gia tăng đáng kể nhờ khả năng thúc đẩy tế bào tua trưởng thành với sự gia tăng biểu hiện của B7-1, B7-2, và CD40. Mặc dù quá trình hoạt hóa tế bào T CD8 + từ con đường tín hiệu của TLR7/TLR8 được kích thích bởi R-848 chưa được hiểu hết, nghiên cứu này cho rằng quá trình trên được đảm nhiệm bởi các tế bào tua biểu hiện CD8 (CD8 + DC) sau khi được kích thích bởi IFN-α được tiết từ các tế bào tua dạng tương bào (plasmacytoid DC/pDC) 100 . Tế bào tua CD8 + , một dạng tế bào tua có nguồn gốc từ dòng lympho, đã được nhiều nghiên cứu chứng minh là có khả năng trình diện chéo kháng nguyên và hoạt hóa tế bào T CD8 + mạnh hơn các tế bào tua từ dòng tủy 101 , 102 , 103 . Nghiên cứu của Cerovic và cộng sự 104 cũng gặt hái được kết quả tương tự khi đánh giá vai trò của tế bào tua CD8 + trong việc hoạt hóa đáp ứng của tế bào T CD8 + ở chuột. Kết quả cho thấy sau khi cảm ứng TLR7/TLR8 bằng R-848, quá trình trình diện chéo kháng nguyên để hoạt hóa tế bào T CD8 + chỉ diễn ra duy nhất ở các tế bào tua CD8 + mà không hề được ghi nhận ở các tế bào tua khác. Những nghiên cứu trên cho thấy các hợp chất IMQ có thể trở thành tá dược miễn dịch tiềm năng để kích thích đáp ứng của tế bào T H 1 lẫn tế bào T CD8 + , mặc dù vai trò và cơ chế trình diện chéo kháng nguyên của các tế bào tua CD8 + ở đây cần được nghiên cứu kĩ hơn.
Tá dược miễn dịch là phối tử của các thụ thể CLR
CLR là các thụ thể lectin thuộc PRR chuyên nhận diện các dạng polysaccharide ở vi khuẩn và nấm. Trong số đó, DEC-205 (CD205) là một thụ thể CLR được biết đến với khả năng kích thích đáp ứng của cả tế bào T H 1 và tế bào T CD8 + . Nghiên cứu của Bonifaz và cộng sự 105 đã thử nghiệm vận chuyển kháng nguyên ovalbumin (OVA) đến tế bào tua bằng cách gắn kết kháng nguyên với kháng thể kháng DEC-205. Kết quả cho thấy OVA sau khi nhập bào đã được tế bào tua trình diện trên MHC-I. Ngoài tế bào T H 1, tế bào T CD8 + đặc hiệu OVA cũng đã được hoạt hóa và hình thành tế bào nhớ với khả năng duy trì đáp ứng sau 90 ngày. Quá trình hoạt hóa tế bào T CD8 + còn được tăng cường khi sử dụng kháng thể kháng CD40 để hỗ trợ tế bào tua trưởng thành. Các nghiên cứu sau đó cũng đạt được kết quả tương đồng khi cho thấy kháng nguyên được vận chuyển bằng kháng thể kháng các CLR khác nhau như langerin, DEC205, và C-type lectin domain family 9 member A (CLEC9A) cũng giúp kích thích đáp ứng của cả tế bào T H 1 và T CD8 + , cũng như khả năng kích thích tế bào tua trình diện chéo kháng nguyên từ con đường tín hiệu của các dạng CLR này cũng đã được ghi nhận, mặc dù cơ chế chưa được khám phá 106 . Một phát hiện khác cho thấy DEC-205 còn có khả năng nhận diện CpG từ đó thúc đẩy đáp ứng của tế bào T CD8 + 107 . Phát hiện này đã bổ sung thêm cho tá dược miễn dịch CpG khả năng kích thích đáp ứng miễn dịch qua trung gian tế bào bằng con đường CLR bên cạnh khả năng kích thích đáp ứng miễn dịch dịch thể thông qua TLR9. Những nghiên cứu trên cho thấy việc nghiên cứu các tá dược nhắm đến CLR (có thể là các phối tử polysaccharide tương ứng hay thậm chí là CpG) có triển vọng phát triển các vaccine gây được đáp ứng miễn dịch của cả tế bào T CD4 + lẫn T CD8 + .
Tá dược miễn dịch kích thích tổn thương mô
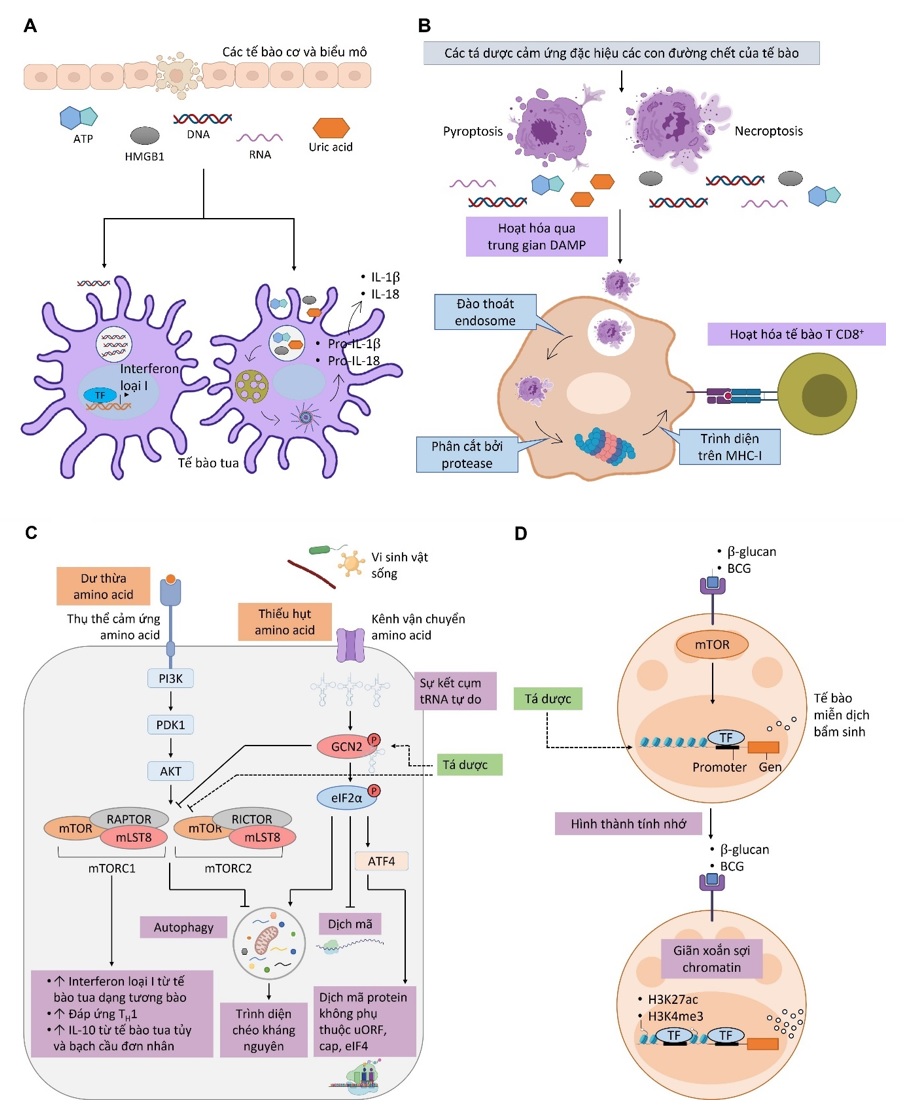
Đến nay, các nghiên cứu đã cho thấy các tế bào trình diện kháng nguyên ngoài khả năng nhận diện PAMP từ vi sinh vật còn có thể nhận diện các DAMP như ATP, uric acid, DNA nội bào, RNA nội bào, hay các protein high mobility group box chromosomal protein 1 (HMGB1) giải phóng từ các tế bào chết, từ đó hoạt hóa hệ miễn dịch thích ứng ( Figure 8 A) 108 , 109 , 110 , 111 . Điển hình cho vai trò của các kiểu mẫu phân tử DAMP trong đáp ứng miễn dịch thích ứng là cơ chế hoạt động của tá dược miễn dịch nhôm. Trong những năm gần đây, các nghiên cứu bắt đầu đặt ra câu hỏi liệu các DAMP có khả năng kích thích đáp ứng miễn dịch qua trung gian tế bào. Từ đây, các nghiên cứu bắt đầu đi sâu hơn để khám phá các cơ chế hoạt hóa hệ miễn dịch thích ứng của DAMP. Kết quả của các nghiên cứu này cho thấy trong một số trường hợp, các DAMP có thể kích thích đáp ứng của tế bào T CD8 + . Thí dụ điển hình nhất là HMGB1, một protein bám DNA giải phóng từ tế bào chết có thể hỗ trợ đáp ứng miễn dịch qua trung gian tế bào. HMGB1 hoạt hóa và kích thích tế bào tua trưởng thành thông qua tương tác với các thụ thể receptor for advanced glycation end product (RAGE) và TLR4 112 , 113 , 114 . Nghiên cứu của Apetoh và cộng sự 115 cho thấy HMGB1 có khả năng thúc đẩy tế bào tua trình diện chéo kháng nguyên thông qua con đường TLR4 để hoạt hóa tế bào T CD8 + . Nghiên cứu lý giải cơ chế này có thể do tín hiệu TLR4 có khả năng ức chế quá trình dung hợp túi phagosome và lysosome đã được chứng minh trong một nghiên cứu trước đó 116 , từ đó tạo cơ hội để kháng nguyên trong phagosome được vận chuyển ra tế bào chất và trình diện trên MHC-I. Một số nghiên cứu khác đã chứng minh HMGB1 có khả năng kích thích quá trình tự thực (autophagy), một quá trình vận chuyển các thành phần trong túi endosome/phagosome ra tế bào chất, ở tế bào ung thư thông qua khả năng tương tác với RAGE cùng với khả năng khóa tương tác giữa Beclin1 và B-cell lymphoma 2 (Bcl-2) 117 , 118 , 119 , 120 . Từ đây, các nghiên cứu đặt ra câu hỏi liệu cơ chế kích thích autophagy này có diễn ra ở tế bào tua để phục vụ cho quá trình trình diện chéo kháng nguyên giúp hoạt hóa tế bào T CD8 + . Tuy nhiên giả thuyết này vẫn chưa được kiểm chứng. Một số nghiên cứu khác về vai trò của các kiểu mẫu phân tử DAMP trong đáp ứng miễn dịch đã cho thấy sự hình thành bẫy ngoại bào bạch cầu trung tính (neutrophil extracellular trap /NET), một dạng bẫy được tiết bởi bạch cầu trung tính bao gồm chromatin và các túi granule dùng để bắt giữ và tiêu diệt vi sinh vật, có thể được kích thích bởi tá dược miễn dịch nhôm 121 , 122 , 123 , và chính NET cũng là nguồn giải phóng HMGB1 bám trên các sợi chromatin 124 giúp hoạt hóa đáp ứng của tế bào T CD8 + trong một số trường hợp. Một kiểu mẫu phân tử DAMP khác là các heat shock protein (Hsp), điển hình là Hsp70. Mặc dù là một chaperone nội bào, Hsp70 lại có vai trò lớn trong việc hoạt hóa hệ miễn dịch bẩm sinh lẫn thích ứng. Nhiều nghiên cứu đã chứng minh Hsp70 kích thích tế bào tua trưởng thành thông qua con đường tín hiệu của TLR2 và TLR4 125 , 126 . Ngoài ra, Hsp70 còn có khả năng kích thích tế bào tua trình diện chéo kháng nguyên và hoạt hóa tế bào T CD8 + 127 , 128 , 129 . Tuy nhiên, các cơ chế tác động này của Hsp70 lên hệ miễn dịch thích ứng vẫn chưa được hiểu rõ. Ngoài ra, một số nghiên cứu cho thấy con đường tín hiệu của STING có thể giúp tế bào tua tăng cường autophagy, trình diện chéo kháng nguyên, và hoạt hóa tế bào T CD8 + 130 , 131 , 132 . Điều này cho thấy việc cảm ứng STING bằng các DNA từ tế bào chết dưới tác động của các tá dược miễn dịch như nhôm có thể giúp kích thích đáp ứng miễn dịch qua trung gian tế bào. Những nghiên cứu trên cho thấy việc phát triển các tá dược miễn dịch mới có khả năng kích thích chọn lọc quá trình giải phóng của các thành phần như DNA nội bào, HMGB1, Hsp70 có thể mở ra bước phát triển cho các thế hệ vaccine mới tạo được đáp ứng miễn dịch qua trung gian tế bào.
Tá dược miễn dịch kích thích apoptosis/necrosis của tế bào
Một số nghiên cứu cho thấy tế bào tua có thể thực bào các tế bào chết và trình diện chéo các kháng nguyên bên trong lên MHC-I. Nghiên cứu của Albert và cộng sự 133 đã thử nghiệm đồng nuôi cấy tế bào tua, tế bào T CD8 + , và bạch cầu đơn nhân nhiễm virus cúm A (influenza A virus). Kết quả cho thấy sau thời gian đồng nuôi cấy, các tế bào tua đã thực bào các bạch cầu bị nhiễm virus đang trong quá trình apoptosis và trình diện chéo các kháng nguyên của virus trên MHC-I cho tế bào T CD8 + nhận diện. Nghiên cứu này đặt ra giả thuyết các tín hiệu apoptosis trong tế bào chết có thể kích thích quá trình trình diện chéo kháng nguyên ở tế bào tua. Giả thuyết này sau đó cũng đã được những nghiên cứu tiếp theo chứng minh là nhờ vào tín hiệu của receptor-interacting protein kinase 1 (RIPK1) và NF-κB trong các tế bào đang ở trạng thái apoptosis và necroptosis 134 . Tá dược miễn dịch MF-59 đã được chứng minh có khả năng kích hoạt quá trình necroptosis của đại thực bào ở hạch để hoạt hóa đáp ứng của tế bào T CD8 + 41 . Những nghiên cứu trên mở ra hướng phát triển các tá dược miễn dịch kích thích quá trình apoptosis của tế bào để tận dụng chính những tế bào chết này làm nguồn cung cấp tín hiệu hỗ trợ quá trình trình diện chéo kháng nguyên của tế bào tua ( Figure 8 B).
Tá dược miễn dịch điều hòa chuyển hóa (metabolic modulator)
Quá trình trình diện chéo kháng nguyên có thể được điều khiển bởi các con đường chuyển hóa bên trong tế bào tua. Cụ thể, trong điều kiện môi trường dư thừa amino acid, các thụ thể amino acid sensing receptor sẽ truyền tín hiệu hoạt hóa các phức hợp mechanistic target of rapamycin (mTOR) làm ức chế quá trình autophagy, từ đó hạn chế việc trình diện chéo các kháng nguyên trong các túi phagosome/endosome. Tuy nhiên, trong điều kiện thiếu nguồn dinh dưỡng từ amino acid, các protein general control nonderepressible 2 kinase (GCN2) được hoạt hóa (bằng cơ chế phosphoryl hóa), sau đó GCN2 tiếp tục phosphoryl hóa nhân tố eukaryotic translation initiation factor 2α (eIF2α) để kích hoạt autophagy giúp các thành phần bên trong các túi phagosome/endosome được chuyển ra tế bào chất để tế bào tận dụng nguồn dinh dưỡng này. Khi đó, các kháng nguyên được chuyển từ túi phagosome/endosome ra tế bào chất có cơ hội được trình diện trên MHC-I ( Figure 8 C) 135 , 16 . Vaccine YF-17D đã được chứng minh có khả năng kích hoạt quá trình trình diện chéo kháng nguyên theo cơ chế trên mặc dù cách thức virus sống trong vaccine hoạt hóa GCN2 vẫn chưa được xác định, tuy nhiên khả năng này lại mất đi khi đánh giá trên virus cùng loại bị bất hoạt bởi tia xạ 135 . Các phát hiện trên mở ra hướng phát triển của các tá dược miễn dịch mới có khả năng kích thích quá trình autophagy của tế bào tua để thúc đẩy trình diện chéo kháng nguyên, thí dụ như các hợp chất giúp kích hoạt quá trình phosphoryl hóa GCN2 hoặc ức chế hình thành các phức hợp mTOR.
Figure 8 . Quá trình kích thích đáp ứng miễn dịch từ mô bị tổn thương (A), tín hiệu apoptosis/necrosis bên trong tế bào (B), quá trình điều hòa chuyển hóa (C), và quá trình điều hòa biểu hiện gene biểu sinh (D) 16
 16" width="300" height="200">
16" width="300" height="200">
CÁC TÁ DƯỢC MIỄN DỊCH KHÁC
Tá dược miễn dịch điều hòa biểu hiện gen biểu sinh (epigenetic modulator)
Bên cạnh tế bào tua và các tế bào của hệ miễn dịch thích ứng, những tế bào khác của hệ miễn dịch bẩm sinh như đại thực bào, bạch cầu đơn nhân, tế bào giết tự nhiên (natural killer cell/NK) cũng đóng vai trò quan trọng và cần được các vaccine nhắm đến. Các nghiên cứu gần đây cho thấy không riêng tế bào của hệ miễn dịch thích ứng, các tế bào của hệ miễn dịch bẩm sinh cũng có tính nhớ đã được ghi nhận trong trường hợp của vaccine Bacillus Calmette-Guérin (BCG) 136 , 137 , 138 , 16 . Không như tính nhớ của tế bào T hay B hình thành từ quá trình nhận diện kháng nguyên và nhân dòng, tính nhớ của tế bào miễn dịch bẩm sinh hình thành do quá trình thay đổi biểu hiện gene biểu sinh (epigenetic change) sau khi tiếp xúc với tác nhân xâm nhiễm 138 , 139 . Sự tái lập trình này giúp hình thành một tính nhớ ngắn hạn và giúp tế bào miễn dịch bẩm sinh hình thành trạng thái “sẵn sàng chiến đấu” trong thời gian từ vài tuần cho đến vài tháng 138 . Trong thời gian này, nếu tiếp xúc lại với tác nhân xâm nhiễm, các tế bào của hệ miễn dịch bẩm sinh sẽ phản ứng nhanh hơn để ngăn chặn tác nhân xâm nhiễm và hỗ trợ hệ miễn dịch thích ứng. Cơ chế này đã được các nghiên cứu chứng minh là do sự tăng cường methyl hóa và acetyl hóa trên các histone H3 ở những vùng gene (trong đó có các gene mã hóa cho các cytokine tiền viêm), giúp các gene này dễ dàng được chuyển sang trạng thái mở để được tăng cường biểu hiện trong những lần tiếp xúc tiếp theo với tác nhân xâm nhiễm ( Figure 8 D) 138 , 139 , 140 , 141 . Một trong số ít các PAMP được chứng minh có khả năng thúc đẩy hình thành tính nhớ ở tế bào miễn dịch bẩm sinh (đại thực bào và bạch cầu đơn nhân) là β-glucan (polysaccharide ở nấm) và LPS thông qua con đường mitogen-activated protein kinase (MAPK) giúp tăng cường quá trình phosphoryl hóa trên cyclic AMP-dependent transcription factor 7 (ATF7), từ đó làm giảm hoạt động của các enzyme H3K9 dimethyltransferase G9a có vai trò co xoắn sợi chromatin được thu hút bởi ATF7, và tăng cường hoạt động của những enzyme như H3K4 methyltransferase giúp sợi chromatin được giãn xoắn và tăng cường biểu hiện gene 142 . Ngoài ra, β-glucan còn có khả năng kích thích quá trình thủy giải đường, có vai trò trong việc hình thành tính nhớ ở đại thực bào và bạch cầu đơn nhân thông qua con đường tín hiệu Akt/mTOR/Hypoxia-inducible factor 1-alpha (HIF1α) 143 , 144 . Những phát hiện trên cho thấy các nghiên cứu sâu hơn về vai trò của những thay đổi biểu hiện gene biểu sinh trong việc hình thành tính nhớ của tế bào miễn dịch bẩm sinh nên được tiến hành để phát triển các tá dược miễn dịch hỗ trợ tương ứng.
Tá dược miễn dịch kích thích tế bào B
Sự biệt hóa của tế bào B thành tương bào tiết kháng thể hoặc tế bào B nhớ trải qua nhiều bước khác nhau trong hạch bạch huyết ( Figure 9 ). Các tế bào T CD4 + sau khi được tế bào tua trình diện kháng nguyên và hoạt hóa ở vùng cận vỏ (paracortex) sẽ di chuyển đến khu vực của tế bào B ở vùng vỏ (cortex) của hạch bạch huyết. Tại đây, tế bào T CD4 + hoạt hóa tế bào B đặc hiệu kháng nguyên nhân dòng 7 , 16 . Các tế bào B này một phần di chuyển đến dây tủy (medullary cord) ở hạch và biệt hóa thành các tương bào sống ngắn (short-lived plasma cell/SLPC) và tiết kháng thể trung hòa vi sinh vật, tuy nhiên nhanh chóng apoptosis ngay sau khi vi sinh vật đã bị loại bỏ 145 , 146 . Số tế bào B còn lại di chuyển về nang tế bào B để hình thành nên trung tâm mầm (germinal center/GC) 7 , 16 , 147 . Một số tế bào T CD4 + sau khi được hoạt hóa tăng biểu hiện các thụ thể C-X-C motif chemokine receptor 5 (CXCR5) và C-C motif chemokine receptor 7 (CCR7) để di chuyển về trung tâm mầm theo tín hiệu hướng hóa của các chemokine đặc trưng ở nang tế bào B và biệt hóa thành tế bào T giúp đỡ ở nang (follicular helper T cell/T FH ) 16 , 147 , 148 , 149 . Tại trung tâm mầm, các tế bào B cạnh tranh lẫn nhau những tín hiệu IL-21 và CD40 ligand (CD40L) (các phân tử kích thích tế bào B ở trung tâm mầm tăng sinh và biệt hóa) từ tế bào T FH phụ thuộc vào mức độ kháng nguyên được tế bào B trình diện trên MHC-II cho tế bào T FH [16]. Bên cạnh đó, thụ thể kháng nguyên của tế bào B trải qua quá trình trưởng thành ái lực (affinity maturation) thông qua sự hình thành số lượng lớn các đột biến sinh dưỡng trên các vùng gene mã hóa của thụ thể. Các tế bào B này cạnh tranh kháng nguyên được trình diện bởi các tế bào tua nang (follicular DC/FDC), từ đó các tế bào B mang thụ thể có ái lực cao với kháng nguyên được chọn lọc và tăng sinh, biệt hóa thành tế bào B nhớ hoặc các tương bào sống lâu (long-lived plasma cell/LLPC), còn các tế bào B mang thụ thể có ái lực thấp với kháng nguyên bị apoptosis do không cạnh tranh được nguồn kháng nguyên với những tế bào B mang thụ thể có ái lực cao 7 , 16 , 150 . Các tế bào B nhớ sau đó có thể tiếp tục ở lại hạch bạch huyết để phản ứng nhanh với các kháng nguyên được chuyển về hạch trong trường hợp tái nhiễm hoặc cũng có thể rời hạch và di chuyển đến máu ngoại vi để “tuần tra” kháng nguyên 151 , 152 , 153 , 154 , 155 , còn các LLPC di chuyển về tủy xương để cư trú, có thể sống rất lâu và tiết kháng thể liên tục trong nhiều năm để bảo vệ cơ thể 16 , 151 , 153 , 156 , 157 . Trong trường hợp tái nhiễm, các tế bào B nhớ nhận diện kháng nguyên nhanh hơn các tế bào B trinh nguyên vì có thụ thể đã trải qua quá trình trưởng thành ái lực, và có khả năng thay thế tế bào tua để trình diện kháng nguyên và hoạt hóa tế bào T H nhớ, cũng như sẵn sàng biệt hóa thành các tương bào tiết kháng thể với số lượng nhiều hơn và ái lực mạnh hơn lần đáp ứng sơ cấp để loại bỏ tác nhân tái xâm nhiễm. Từ đây, có thể thấy các tế bào B nhớ và LLPC có vai trò quan trọng trong việc bảo vệ cơ thể trong thời gian dài kể cả khi tác nhân xâm nhiễm đã bị loại bỏ. Điều này cho thấy tá dược miễn dịch tác động lên tế bào B ở trung tâm mầm và tế bào tua nang cần được phát triển để kích thích quá trình hình thành tế bào B nhớ và LLPC. Một trong các tá dược miễn dịch có khả năng trên là 3M-052 với khả năng thúc đẩy tế bào LLPC đặc hiệu kháng nguyên HIV-1 hình thành và tồn tại trong thời gian 70 tuần trong tủy xương 158 .
Figure 9 . Quá trình hình thành tế bào B nhớ và LLPC 16
 16" width="300" height="200">
16" width="300" height="200">
LỰA CHỌN TÁ DƯỢC MIỄN DỊCH CHO VACCINE COVID-19 VÀ NHỮNG THÁCH THỨC
Một trong những tiêu chí quan trọng nhất để đánh giá hiệu quả của vaccine COVID-19 là khả năng thúc đẩy cơ thể tạo kháng thể trung hòa virus SARS-CoV-2. Ngoài ra, sự hình thành các tế bào nhớ cũng rất quan trọng để bảo vệ cơ thể trong thời gian dài. Sự hình thành các tế bào T nhớ cư trú trong một thời gian dài ở mô (tissue-resident memory T cell/T RM ) là rất cần thiết để “trực chiến” nhằm nhanh chóng phối hợp với các tế bào của hệ miễn dịch bẩm sinh kích hoạt trạng thái “sẵn sàng chiến đấu” nếu phát hiện có vi sinh vật tái nhiễm. Không những ở mô, dòng máu trong cơ thể cũng cần được thiết lập tuyến phòng thủ bởi những tế bào B nhớ. Ngoài ra, do virus là vật thể sống phụ thuộc vào quá trình tăng trưởng nội bào, bên cạnh đáp ứng tạo kháng thể trung hòa virus, đáp ứng của tế bào T CD8 + cũng cần được kích hoạt để tiêu diệt các tế bào nhiễm là nơi ẩn náu và sao chép của virus. Tuy nhiên, không phải vaccine COVID-19 nào cũng đáp ứng đủ tất cả các tiêu chí trên vì tính phụ thuộc vào thành phần tá dược miễn dịch sử dụng. Đứng trước tình trạng khẩn cấp khi SARS-CoV-2 đã lây lan trên toàn cầu, việc chờ đợi sự ra đời của những tá dược miễn dịch mới giúp vaccine đáp ứng được toàn bộ những yêu cầu trên là không thể. Thay vào đó, các vaccine cần phải tận dụng những tá dược miễn dịch đã có sẵn và đã được cấp phép sử dụng trên người một cách hiệu quả nhất. Vì vậy, việc lựa chọn một tá dược miễn dịch phù hợp dựa trên sự so sánh ưu nhược điểm của các tá dược miễn dịch đã được cấp phép ( Table 1 ) trực tiếp quyết định đến hiệu quả của các vaccine COVID-19 ở thời điểm hiện tại.
Hiện nay, tương đối nhiều vaccine COVID-19 sử dụng tá dược miễn dịch nhôm (Vero Cell, CoronaVac, MVC-COV1901, Nanocovax) và tá dược miễn dịch nhũ hóa (vaccine của Safoni/GSK hay Medicago) nhờ khả năng kích thích được đáp ứng miễn dịch dịch thể lẫn đáp ứng miễn dịch qua trung gian tế bào của hai loại tá dược miễn dịch này, mặc dù một số vaccine trong số này vẫn còn trong giai đoạn thử nghiệm lâm sàng 2 , 16 . Mặc dù vậy, liều lượng của những tá dược miễn dịch này cũng cần được tối ưu hóa để hạn chế tối đa tác dụng phụ. Hơn nữa, vấn đề đáp ứng miễn dịch theo hướng T H 2 của các vaccine sử dụng tá dược miễn dịch nhôm cũng cần được xem xét kĩ khi một số vaccine như Vero Cell đã ghi nhận phản ứng dị ứng (nổi mẫn đỏ) ở một số trường hợp 159 . Bên cạnh đó, tuy các tá dược miễn dịch nhôm và chất nhũ hóa có thể kích thích đáp ứng miễn dịch qua trung gian tế bào, các đánh giá về đáp ứng của tế bào T CD8 + cũng cần được đánh giá ở những vaccine sử dụng hai dạng tá dược miễn dịch này. Ngoài ra, việc phối hợp nhiều tá dược miễn dịch khác nhau, như phối tử của TLR7/TLR8 kết hợp nhôm trong vaccine Covaxin (BBV152) của Bharath Biotech 160 hay CpG 1018 kết hợp nhôm trong vaccine MVC-COV1901 24 , cũng nên được cân nhắc áp dụng để điều hướng đáp ứng miễn dịch theo những hướng mong muốn.
Hiện nay, một số vaccine đặc biệt như Vaxzevria, Comirnaty, Spikevax được sản xuất không cần tá dược miễn dịch bởi chính thành phần gây đáp ứng của các vaccine này đã mang đặc tính của tá dược miễn dịch. Cụ thể, vaccine Vaxzevria sử dụng vector adenovirus được chuyển gene mã hóa kháng nguyên của SARS-CoV-2 để tế bào trong cơ thể biểu hiện ra kháng nguyên virus trên bề mặt 3 , 161 , 162 , do đó thành phần DNA virus đã đóng vai trò là tá dược miễn dịch cảm ứng các thụ thể PRR nhận diện nucleic acid, thí dụ như cGAS cảm ứng con đường STING ở tế bào tua 163 , 164 , 165 . Tương tự, hai vaccine Comirnaty và Spikevax sử dụng mRNA vừa giữ nhiệm vụ mã hóa cho kháng nguyên của SARS-CoV-2 vừa đóng vai trò là tá dược miễn dịch theo cơ chế trên 166 , 167 , 168 , 169 . Hai vaccine mRNA này còn được chứng minh là có khả năng kích hoạt đáp ứng của tế bào T CD8 + trong cơ thể 170 , 171 , mặc dù cơ chế ở đây chưa được chứng minh. Khả năng này đã bổ sung thêm cho các vaccine mRNA một điểm cộng bên cạnh tính an toàn cao. Một số nghiên cứu cho rằng khả năng hoạt hóa tế bào T CD8 + của hai vaccine trên là nhờ thành phần mRNA trong vaccine có khả năng cảm ứng các thụ thể PRR nhận diện RNA giúp kích hoạt một số con đường tín hiệu chưa được biết đến trong tế bào tua góp phần trong quá trình hoạt hóa tế bào T CD8 + 16 . Ngoài ra, các nghiên cứu cũng đã chứng minh ngoài việc mRNA được vận chuyển (bằng các hạt liposome) đến các tế bào trong mô tại vị trí gây đáp ứng để các tế bào này biểu hiện ra kháng nguyên virus ở dạng tiết hoặc đính màng, các mRNA còn có thể được đưa trực tiếp đến tế bào tua 172 . Với bản chất là liposome tích điện dương ở pH acid, các hạt liposome này sau khi được thực bào/nhập bào trong túi phagosome/endosome có tính acid dung hợp với màng phospholipid tích điện âm của phagosome/endosome 173 , 174 , từ đó mRNA được chuyển ra tế bào chất và được tế bào tua dịch mã ra protein kháng nguyên của virus, sau đó được trình diện trên MHC-I để hoạt hóa tế bào T CD8 + .
Từ câu chuyện của vaccine sốt vàng da YF-17D sử dụng miễn dịch nguyên là virus sống có khả năng sao chép trong cơ thể, một câu hỏi được dấy lên là liệu các vaccine cung cấp được nguồn kháng nguyên liên tục trong thời gian dài (như virus sống) có thể kích hoạt được đáp ứng miễn dịch qua trung gian tế bào hay không 16 . Giả thuyết này bắt đầu được củng cố hơn khi đáp ứng của tế bào T CD8 + cũng được ghi nhận ở các vaccine Comirnaty hay Spikevax vốn dĩ sử dụng mRNA có thể giúp tế bào trong mô duy trì biểu hiện kháng nguyên trong một thời gian nhất định 16 . Hiện nay, một số vaccine COVID-19 cũng sử dụng miễn dịch nguyên là hạt virus nhưng đều là virus không có khả năng tăng trưởng trong cơ thể do bị bất hoạt bằng tác nhân nhiệt, hóa chất, tia xạ (Vero Cell, CoronaVac) hoặc bị biến đổi di truyền để mất khả năng sao chép (Vaxzevria). Từ đây, có thể đặt ra câu hỏi nếu các vaccine trên chỉ sử dụng virus nhược độc (bằng cách nuôi cấy lặp lại nhiều lần trên dòng tế bào Vero để làm giảm độc lực nhưng vẫn duy trì được khả năng sống) thì liệu có khả năng kích thích đáp ứng của tế bào T CD8 + như cách vaccine YF-17D đã làm được (bằng cách duy trì nguồn kháng nguyên trong một khoảng thời gian và kích thích quá trình autophagy để tế bào tua trình diện chéo kháng nguyên). Tuy nhiên, chiến lược sử dụng virus nhược độc luôn khá mạo hiểm khi chứa đựng nguy cơ hồi tính của virus gây đe dọa tính mạng.
KẾT LUẬN
Nhìn chung, các vaccine hiện nay vẫn chỉ đang chủ yếu sử dụng bốn nhóm tá dược miễn dịch chính là nhôm, chất nhũ hóa, LPS, và CpG, mặc dù còn rất nhiều các thụ thể nhận diện kiểu mẫu phân tử ở tế bào trình diện kháng nguyên vẫn chưa được khai thác. Tóm lại, đại dịch COVID-19 cho chúng ta cơ hội nhìn nhận rằng cho đến nay, con người vẫn đang có rất ít các tá dược miễn dịch cho vaccine. Điều này đặt ra một yêu cầu cấp thiết trong việc phát triển các thế hệ tá dược miễn dịch mới (đặc biệt là các tá dược miễn dịch kích thích được cả đáp ứng của tế bào TH1 lẫn tế bào T CD8+ cùng với khả năng thúc đẩy mạnh mẽ hệ miễn dịch hình thành các tế bào nhớ) để phục vụ cho việc sản xuất vaccine giúp con người đối mặt với thách thức từ các dịch bệnh khác trong tương lai.
DANH MỤC CÁC TỪ VIẾT TẮT
3D-PHAD: 3-Deacylphosphorylated hexaacyl disaccharide
ACE2: Angiotensin-converting enzyme 2
AS: Adjuvant system
ATF7: Cyclic AMP-dependent transcription factor 7
ATP: Adenosine triphosphate
BCG: Bacillus Calmette-Guérin
Bcl-2: B-cell lymphoma 2
CCL: C-C motif chemokine ligand
CCR7: C-C motif chemokine receptor 7
CD: Cluster of differentiation
cGAS: Cyclic guanosine monophosphate-adenosine monophosphate synthase
CLEC9A: C-type lectin domain family 9 member A
CLR: C-type lectin receptor
COVID-19: Coronavirus disease 2019
CpG: Cytosine-phosphate-guanine
CXCL: C-X-C motif chemokine ligand
CXCR5: C-X-C motif chemokine receptor 5
DAMP: Damage-associated molecular pattern
DC: Dendritic cell
DNA: Deoxyribonucleic acid
dsRNA: Double-stranded ribonucleic acid
eIF2α: Eukaryotic translation initiation factor 2α
ERAP2: Endoplasmic reticulum aminopeptidase 2
FDC: Follicular dendritic cell
GC: Germinal center
GCN2: General control nonderepressible 2 kinase
G-CSF: Granulocyte colony-stimulating factor
GpC: Guanine-phosphate-cytosine
HIF1α: Hypoxia-inducible factor 1-alpha
HIV-1: Human immunodeficiency virus 1
HMGB1: High mobility group box chromosomal protein 1
Hsp: Heat shock protein
IFN: Interferon
IL: Interleukin
IMQ: Imidazoquinoline
IRF: Interferon regulatory factor
JNK1: c-Jun N-terminal kinase 1
LAMP-1: Lysosome-associated membrane protein-1
LLPC: Long-lived plasma cell
LPS: Lipopolysaccharide
MAPK: Mitogen-activated protein kinase
MDA5: Melanoma differentiation-associated protein 5
MHC: Major histocompatibility complex
MPL: Monophosphoryl lipid A
MyD88: Myeloid differentiation factor 88
mRNA: Messenger ribonucleic acid
mTOR: Mechanistic target of rapamycin
NET: Neutrophil extracellular trap
NF-κB: Nuclear factor κB
NK: Natural killer cell
NLRP3: NACHT, LRR and PYD domains-containing protein 3
ODN: Oligodeoxynucleotide
OVA: Ovalbumin
PAMP: Pathogen-associated molecular pattern
pDC: Plasmacytoid dendritic cell
Poly(I:C); Polyinosinic-polycytidylic acid
PRR: Pattern recognition receptor
RAGE: Receptor for advanced glycation end product
RIG-I: Retinoic acid-inducible gene I
RIPK1: Receptor-interacting protein kinase 1
RNA: Ribonucleic acid
SARS-CoV-2: Severe acute respiratory syndrome coronavirus 2
SLPC: Short-lived plasma cell
ssRNA: Single-stranded ribonucleic acid
STING: Stimulator of interferon gene
Syk: Spleen tyrosine kinase
TCR: T-cell receptor
TFH: Follicular helper T cell
TfR: Transferrin receptor
TH: Helper T cell
TLR: Toll-like receptor
TNF: Tumor necrosis factor
TPP2: Tripeptidyl-peptidase 2
TRIF: TIR-domain-containing adapter-inducing interferon-β
TRM: Tissue-resident memory T cell
YF-17D: Yellow fever 17D
XUNG ĐỘT LỢI ÍCH
Các tác giả cam kết không có xung đột lợi ích.
ĐÓNG GÓP CỦA CÁC TÁC GIẢ
Các tác giả Lê Khánh Thiên, Phạm Hoàng Tính tham gia viết bản thảo.
Tác giả Trần Văn Hiếu tham gia chỉnh sửa bản thảo.
Tất cả tác giả đồng ý với bản cuối cùng của bản thảo.
References
- Awadasseid A, Wu Y, Tanaka Y, Zhang W. Current advances in the development of SARS-CoV-2 vaccines. Int J Biol Sci. 2021;17(1):8-19. . ;:. PubMed Google Scholar
- Palacios R, Patino EG, de Oliveira Piorelli R, Conde M, Batista AP, Zeng G, Double-blind, randomized, placebo-controlled phase III clinical trial to evaluate the efficacy and safety of treating healthcare professionals with the adsorbed COVID-19 (Inactivated) Vaccine Manufactured by Sinovac - PROFISCOV: A structured summary of a study protocol for a randomised controlled trial. Trials. 2020;21(1):853. . ;:. PubMed Google Scholar
- Soiza RL, Scicluna C, Thomson EC. Efficacy and safety of COVID-19 vaccines in older people. Age Ageing. 2021;50(2):279-83. . ;:. PubMed Google Scholar
- Keech C, Albert G, Cho I, Robertson A, Reed P, Neal S,. Phase 1-2 Trial of a SARS-CoV-2 Recombinant spike protein nanoparticle vaccine. N Engl J Med. 2020;383(24):2320-32. . ;:. PubMed Google Scholar
- Chung YH, Beiss V, Fiering SN, Steinmetz NF. COVID-19 Vaccine frontrunners and their nanotechnology design. ACS Nano. 2020;14(10):12522-37. . ;:. PubMed Google Scholar
- Ali A, Vijayan R. Dynamics of the ACE2-SARS-CoV-2/SARS-CoV spike protein interface reveal unique mechanisms. Sci Rep. 2020;10(1):14214. . ;:. PubMed Google Scholar
- Hiếu TV. Miễn dịch học: Chức năng và các bất thường của hệ miễn dịch. Thành phố Hồ Chí Minh, Việt Nam: NXB Đại học Quốc gia Thành phố Hồ Chí Minh; 2016. . ;:. Google Scholar
- Sharpe AH, Freeman GJ. The B7-CD28 superfamily. Nat Rev Immunol. 2002;2(2):116-26. . ;:. PubMed Google Scholar
- Riella LV, Sayegh MH. T-cell co-stimulatory blockade in transplantation: two steps forward one step back! Expert Opin Biol Ther. 2013;13(11):1557-68. . ;:. PubMed Google Scholar
- Ippoliti G, D'Armini AM, Lucioni M, Marjieh M, Vigano M. Introduction to the use of belatacept: a fusion protein for the prevention of posttransplant kidney rejection. Biologics. 2012;6:355-62. . ;:. PubMed Google Scholar
- Kawai T, Akira S. The role of pattern-recognition receptors in innate immunity: update on Toll-like receptors. Nat Immunol. 2010;11(5):373-84. . ;:. PubMed Google Scholar
- Mogensen TH. Pathogen recognition and inflammatory signaling in innate immune defenses. Clin Microbiol Rev. 2009;22(2):240-73, Table of Contents. . ;:. PubMed Google Scholar
- Medzhitov R. Toll-like receptors and innate immunity. Nat Rev Immunol. 2001;1(2):135-45. . ;:. PubMed Google Scholar
- Geijtenbeek TB, Gringhuis SI. Signalling through C-type lectin receptors: shaping immune responses. Nat Rev Immunol. 2009;9(7):465-79. . ;:. PubMed Google Scholar
- Honda K, Taniguchi T. IRFs: master regulators of signalling by Toll-like receptors and cytosolic pattern-recognition receptors. Nat Rev Immunol. 2006;6(9):644-58. . ;:. PubMed Google Scholar
- Pulendran B, P SA, O'Hagan DT. Emerging concepts in the science of vaccine adjuvants. Nat Rev Drug Discov. 2021;20(6):454-75. . ;:. PubMed Google Scholar
- Doyle SE, Vaidya SA, O'Connell R, Dadgostar H, Dempsey PW, Wu T-T,. IRF3 Mediates a TLR3/TLR4-specific antiviral gene program. Immunity. 2002;17(3):251-63. . ;:. Google Scholar
- Wu J, Chen ZJ. Innate immune sensing and signaling of cytosolic nucleic acids. Annu Rev Immunol. 2014;32:461-88. . ;:. PubMed Google Scholar
- Bermejo-Jambrina M, Eder J, Helgers LC, Hertoghs N, Nijmeijer BM, Stunnenberg M. C-type lectin receptors in antiviral immunity and viral escape. Front Immunol. 2018;9:590. . ;:. PubMed Google Scholar
- Dalod M, Chelbi R, Malissen B, Lawrence T. Dendritic cell maturation: functional specialization through signaling specificity and transcriptional programming. EMBO J. 2014;33(10):1104-16. . ;:. PubMed Google Scholar
- Bai S, Dong A. Effects of immobilization onto aluminum hydroxide particles on the thermally induced conformational behavior of three model proteins. Int J Biol Macromol. 2009;45(1):80-5. . ;:. PubMed Google Scholar
- Tseng CT, Sbrana E, Iwata-Yoshikawa N, Newman PC, Garron T, Atmar RL,. Immunization with SARS coronavirus vaccines leads to pulmonary immunopathology on challenge with the SARS virus. PLoS One. 2012;7(4):e35421. . ;:. PubMed Google Scholar
- Wang L, Shi W, Joyce MG, Modjarrad K, Zhang Y, Leung K,. Evaluation of candidate vaccine approaches for MERS-CoV. Nat Commun. 2015;6:7712. . ;:. PubMed Google Scholar
- Hsieh SM, Liu WD, Huang YS, Lin YJ, Hsieh EF, Lian WC, Safety and immunogenicity of a recombinant stabilized prefusion SARS-CoV-2 spike protein vaccine (MVC-COV1901) adjuvanted with CpG 1018 and aluminum hydroxide in healthy adults: A Phase 1, dose-escalation study. EClinicalMedicine. 2021;38:100989. . ;:. Google Scholar
- Hutchison S, Benson RA, Gibson VB, Pollock AH, Garside P, Brewer JM. Antigen depot is not required for alum adjuvanticity. FASEB J. 2012;26(3):1272-9. . ;:. PubMed Google Scholar
- Marichal T, Ohata K, Bedoret D, Mesnil C, Sabatel C, Kobiyama K, DNA released from dying host cells mediates aluminum adjuvant activity. Nat Med. 2011;17(8):996-1002. . ;:. Google Scholar
- McKee AS, Burchill MA, Munks MW, Jin L, Kappler JW, Friedman RS, Host DNA released in response to aluminum adjuvant enhances MHC class II-mediated antigen presentation and prolongs CD4 T-cell interactions with dendritic cells. Proc Natl Acad Sci U S A. 2013;110(12):E1122-31. . ;:. PubMed Google Scholar
- Riteau N, Baron L, Villeret B, Guillou N, Savigny F, Ryffel B, ATP release and purinergic signaling: a common pathway for particle-mediated inflammasome activation. Cell Death Dis. 2012;3:e403. . ;:. Google Scholar
- Kool M, Soullie T, van Nimwegen M, Willart MA, Muskens F, Jung S, Alum adjuvant boosts adaptive immunity by inducing uric acid and activating inflammatory dendritic cells. J Exp Med. 2008;205(4):869-82. . ;:. Google Scholar
- Kool M, Petrilli V, De Smedt T, Rolaz A, Hammad H, van Nimwegen M, Cutting edge: alum adjuvant stimulates inflammatory dendritic cells through activation of the NALP3 inflammasome. J Immunol. 2008;181(6):3755-9. . ;:. PubMed Google Scholar
- Eisenbarth SC, Colegio OR, O'Connor W, Sutterwala FS, Flavell RA. Crucial role for the Nalp3 inflammasome in the immunostimulatory properties of aluminium adjuvants. Nature. 2008;453(7198):1122-6. . ;:. Google Scholar
- Winter WE, Hardt NS, Fuhrman S. Immunoglobulin E: importance in parasitic infections and hypersensitivity responses. Arch Pathol Lab Med. 2000;124(9):1382-5. . ;:. PubMed Google Scholar
- Batista-Duharte A, Martinez DT, Carlos IZ. Efficacy and safety of immunological adjuvants. Where is the cut-off? Biomed Pharmacother. 2018;105:616-24. . ;:. PubMed Google Scholar
- Diehl S, Anguita J, Hoffmeyer A, Zapton T, Ihle JN, Fikrig E, Inhibition of Th1 differentiation by IL-6 is mediated by SOCS1. Immunity. 2000;13(6):805-15. . ;:. Google Scholar
- Diehl S, Rincón M. The two faces of IL-6 on Th1/Th2 differentiation. Molecular Immunology. 2002;39(9):531-6. . ;:. Google Scholar
- Choi P, Reiser H. IL-4: role in disease and regulation of production. Clin Exp Immunol. 1998;113(3):317-9. . ;:. PubMed Google Scholar
- Yoshimoto T. The Hunt for the source of primary interleukin-4: How we discovered that natural killer T cells and Basophils determine T helper type 2 cell differentiation in vivo. Front Immunol. 2018;9:716. . ;:. Google Scholar
- Punnonen J, Yssel H, Devries J. The relative contribution of IL-4 and IL-13 to human IgE synthesis induced by activated CD4 or CD8 T cells. Journal of Allergy and Clinical Immunology. 1997;100(6):792-801. . ;:. Google Scholar
- Pene J, Rousset F, Briere F, Chretien I, Bonnefoy JY, Spits H. IgE production by normal human lymphocytes is induced by interleukin 4 and suppressed by interferons gamma and alpha and prostaglandin E2. Proc Natl Acad Sci U S A. 1988;85(18):6880-4. . ;:. PubMed Google Scholar
- Igbokwe IO, Igwenagu E, Igbokwe NA. Aluminium toxicosis: a review of toxic actions and effects. Interdiscip Toxicol. 2019;12(2):45-70. . ;:. PubMed Google Scholar
- Kim EH, Woodruff MC, Grigoryan L, Maier B, Lee SH, Mandal P, et al. Squalene emulsion-based vaccine adjuvants stimulate CD8 T cell, but not antibody responses, through a RIPK3-dependent pathway. Elife. 2020;9. . ;:. PubMed Google Scholar
- Allison AC. Squalene and squalane emulsions as adjuvants. Methods. 1999;19(1):87-93. . ;:. PubMed Google Scholar
- O'Hagan DT, Ott GS, Nest GV, Rappuoli R, Giudice GD. The history of MF59((R)) adjuvant: a phoenix that arose from the ashes. Expert Rev Vaccines. 2013;12(1):13-30. . ;:. PubMed Google Scholar
- Calabro S, Tortoli M, Baudner BC, Pacitto A, Cortese M, O'Hagan DT. Vaccine adjuvants alum and MF59 induce rapid recruitment of neutrophils and monocytes that participate in antigen transport to draining lymph nodes. Vaccine. 2011;29(9):1812-23. . ;:. PubMed Google Scholar
- Seubert A, Monaci E, Pizza M, O'Hagan DT, Wack A. The adjuvants aluminum hydroxide and MF59 induce monocyte and granulocyte chemoattractants and enhance monocyte differentiation toward dendritic cells. J Immunol. 2008;180(8):5402-12. . ;:. PubMed Google Scholar
- Vono M, Taccone M, Caccin P, Gallotta M, Donvito G, Falzoni S. The adjuvant MF59 induces ATP release from muscle that potentiates response to vaccination. Proc Natl Acad Sci U S A. 2013;110(52):21095-100. . ;:. PubMed Google Scholar
- Ko EJ, Lee YT, Kim KH, Jung YJ, Lee Y, Denning TL, Effects of MF59 adjuvant on induction of isotype-Switched IgG antibodies and protection after immunization with T-dependent Influenza virus vaccine in the absence of CD4+ T cells. J Virol. 2016;90(15):6976-88. . ;:. PubMed Google Scholar
- Morel S, Didierlaurent A, Bourguignon P, Delhaye S, Baras B, Jacob V, Adjuvant System AS03 containing alpha-tocopherol modulates innate immune response and leads to improved adaptive immunity. Vaccine. 2011;29(13):2461-73. . ;:. PubMed Google Scholar
- Howard LM, Hoek KL, Goll JB, Samir P, Galassie A, Allos TM. Cell-based systems biology analysis of human AS03-adjuvanted H5N1 Avian influenza vaccine responses: A Phase I Randomized Controlled Trial. PLoS One. 2017;12(1):e0167488. . ;:. PubMed Google Scholar
- Lu YC, Yeh WC, Ohashi PS. LPS/TLR4 signal transduction pathway. Cytokine. 2008;42(2):145-51. PMid:. ;:. PubMed Google Scholar
- Schmittel A, Scheibenbogen C, Keilholz U. Lipopolysaccharide effectively up-regulates B7-1 (CD80) expression and costimulatory function of human monocytes. Scand J Immunol. 1995;42(6):701-4. . ;:. Google Scholar
- Watanabe S, Inoue J. Intracellular delivery of lipopolysaccharide induces effective Th1-immune responses independent of IL-12. PLoS One. 2013;8(7):e68671. . ;:. PubMed Google Scholar
- Yamamoto Y, Harashima A, Saito H, Tsuneyama K, Munesue S, Motoyoshi S,. Septic shock is associated with receptor for advanced glycation end products ligation of LPS. J Immunol. 2011;186(5):3248-57. . ;:. PubMed Google Scholar
- Johnson AG, Tomai M, Solem L, Beck L, Ribi E. Characterization of a nontoxic monophosphoryl lipid A. Rev Infect Dis. 1987;9 Suppl 5:S512-6. . ;:. PubMed Google Scholar
- Casella CR, Mitchell TC. Putting endotoxin to work for us: monophosphoryl lipid A as a safe and effective vaccine adjuvant. Cell Mol Life Sci. 2008;65(20):3231-40. . ;:. PubMed Google Scholar
- Johnson DA, Keegan DS, Sowell CG, Livesay MT, Johnson CL, Taubner LM, 3-O-Desacyl monophosphoryl lipid A derivatives: synthesis and immunostimulant activities. J Med Chem. 1999;42(22):4640-9. . ;:. PubMed Google Scholar
- Cekic C, Casella CR, Eaves CA, Matsuzawa A, Ichijo H, Mitchell TC. Selective activation of the p38 MAPK pathway by synthetic monophosphoryl lipid A. J Biol Chem. 2009;284(46):31982-91. . ;:. PubMed Google Scholar
- Thompson BS, Chilton PM, Ward JR, Evans JT, Mitchell TC. The low-toxicity versions of LPS, MPL adjuvant and RC529, are efficient adjuvants for CD4+ T cells. J Leukoc Biol. 2005;78(6):1273-80. . ;:. PubMed Google Scholar
- Mata-Haro V, Cekic C, Martin M, Chilton PM, Casella CR, Mitchell TC. The vaccine adjuvant monophosphoryl lipid A as a TRIF-biased agonist of TLR4. Science. 2007;316(5831):1628-32. . ;:. PubMed Google Scholar
- Didierlaurent AM, Morel S, Lockman L, Giannini SL, Bisteau M, Carlsen H, AS04, an aluminum salt- and TLR4 agonist-based adjuvant system, induces a transient localized innate immune response leading to enhanced adaptive immunity. J Immunol. 2009;183(10):6186-97. . ;:. PubMed Google Scholar
- Garcon N, Morel S, Didierlaurent A, Descamps D, Wettendorff M, Van Mechelen M. Development of an AS04-adjuvanted HPV vaccine with the adjuvant system approach. BioDrugs. 2011;25(4):217-26. . ;:. PubMed Google Scholar
- Coccia M, Collignon C, Herve C, Chalon A, Welsby I, Detienne S, Cellular and molecular synergy in AS01-adjuvanted vaccines results in an early IFNgamma response promoting vaccine immunogenicity. NPJ Vaccines. 2017;2:25. . ;:. PubMed Google Scholar
- Didierlaurent AM, Laupeze B, Di Pasquale A, Hergli N, Collignon C, Garcon N. Adjuvant system AS01: helping to overcome the challenges of modern vaccines. Expert Rev Vaccines. 2017;16(1):55-63. . ;:. PubMed Google Scholar
- Welsby I, Detienne S, N'Kuli F, Thomas S, Wouters S, Bechtold V,. Lysosome-dependent activation of human dendritic cells by the vaccine adjuvant QS-21. Front Immunol. 2016;7:663. . ;:. PubMed Google Scholar
- Bode C, Zhao G, Steinhagen F, Kinjo T, Klinman DM. CpG DNA as a vaccine adjuvant. Expert Rev Vaccines. 2011;10(4):499-511. . ;:. PubMed Google Scholar
- Campbell JD. Development of the CpG Adjuvant 1018: A case study. Methods Mol Biol. 2017;1494:15-27. . ;:. PubMed Google Scholar
- Dalpke AH, Zimmermann S, Albrecht I, Heeg K. Phosphodiester CpG oligonucleotides as adjuvants: polyguanosine runs enhance cellular uptake and improve immunostimulative activity of phosphodiester CpG oligonucleotides in vitro and in vivo. Immunology. 2002;106(1):102-12. . ;:. PubMed Google Scholar
- Chen W, Jiang M, Yu W, Xu Z, Liu X, Jia Q. CpG-Based nanovaccines for cancer immunotherapy. Int J Nanomedicine. 2021;16:5281-99. . ;:. PubMed Google Scholar
- Guiducci C, Ott G, Chan JH, Damon E, Calacsan C, Matray T. Properties regulating the nature of the plasmacytoid dendritic cell response to Toll-like receptor 9 activation. J Exp Med. 2006;203(8):1999-2008. . ;:. PubMed Google Scholar
- Kuo TY, Lin MY, Coffman RL, Campbell JD, Traquina P, Lin YJ, Development of CpG-adjuvanted stable prefusion SARS-CoV-2 spike antigen as a subunit vaccine against COVID-19. Sci Rep. 2020;10(1):20085. . ;:. PubMed Google Scholar
- Newton SM, Wasley RD, Wilson A, Rosenberg LT, Miller JF, Stocker BA. Segment IV of a Salmonella flagellin gene specifies flagellar antigen epitopes. Mol Microbiol. 1991;5(2):419-25. . ;:. PubMed Google Scholar
- Hajam IA, Dar PA, Shahnawaz I, Jaume JC, Lee JH. Bacterial flagellin-a potent immunomodulatory agent. Exp Mol Med. 2017;49(9):e373. . ;:. PubMed Google Scholar
- Vijay-Kumar M, Aitken JD, Sanders CJ, Frias A, Sloane VM, Xu J, Flagellin treatment protects against chemicals, bacteria, viruses, and radiation. J Immunol. 2008;180(12):8280-5. . ;:. PubMed Google Scholar
- Honko AN, Mizel SB. Mucosal administration of flagellin induces innate immunity in the mouse lung. Infect Immun. 2004;72(11):6676-9. . ;:. PubMed Google Scholar
- Mizel SB, Honko AN, Moors MA, Smith PS, West AP. Induction of macrophage nitric oxide production by Gram-negative flagellin involves signaling via heteromeric Toll-like receptor 5/Toll-like receptor 4 complexes. J Immunol. 2003;170(12):6217-23. . ;:. PubMed Google Scholar
- Means TK, Hayashi F, Smith KD, Aderem A, Luster AD. The Toll-like receptor 5 stimulus bacterial flagellin induces maturation and chemokine production in human dendritic cells. J Immunol. 2003;170(10):5165-75. . ;:. PubMed Google Scholar
- Tran HV, Nguyen AV, Tran CTB. Cloning, expression and purification of the recombinant FliC from Salmonella enteritidis. Science and Technology Development Journal. 2016;19(4):62-9. . ;:. Google Scholar
- Tran B-CT, Vo-Nguyen H-V, Nguyen V-A, Tran TL, Tran-Van H. FliC-delta220-320 from Salmonella enteritidis as an adjuvant for vaccine development. SSR Institute of International Journal of Life Sciences. 2020;6(2):2494-501. . ;:. Google Scholar
- Ho NI, Huis In 't Veld LGM, Raaijmakers TK, Adema GJ. Adjuvants enhancing cross-presentation by dendritic cells: The key to more effective vaccines? Front Immunol. 2018;9:2874. . ;:. PubMed Google Scholar
- Fehres CM, Unger WW, Garcia-Vallejo JJ, van Kooyk Y. Understanding the biology of antigen cross-presentation for the design of vaccines against cancer. Front Immunol. 2014;5:149. . ;:. PubMed Google Scholar
- Du G, Sun X. Engineering nanoparticulate vaccines for enhancing antigen cross-presentation. Curr Opin Biotechnol. 2020;66:113-22. . ;:. PubMed Google Scholar
- Akondy RS, Johnson PL, Nakaya HI, Edupuganti S, Mulligan MJ, Lawson B. Initial viral load determines the magnitude of the human CD8 T cell response to yellow fever vaccination. Proc Natl Acad Sci USA. 2015;112(10):3050-5. . ;:. PubMed Google Scholar
- Fortier ME, Kent S, Ashdown H, Poole S, Boksa P, Luheshi GN. The viral mimic, polyinosinic:polycytidylic acid, induces fever in rats via an interleukin-1-dependent mechanism. Am J Physiol Regul Integr Comp Physiol. 2004;287(4):R759-66. . ;:. PubMed Google Scholar
- Hafner AM, Corthesy B, Merkle HP. Particulate formulations for the delivery of poly(I:C) as vaccine adjuvant. Adv Drug Deliv Rev. 2013;65(10):1386-99. . ;:. PubMed Google Scholar
- Tewari K, Flynn BJ, Boscardin SB, Kastenmueller K, Salazar AM, Anderson CA,. Poly(I:C) is an effective adjuvant for antibody and multi-functional CD4+ T cell responses to Plasmodium falciparum circumsporozoite protein (CSP) and alphaDEC-CSP in non human primates. Vaccine. 2010;28(45):7256-66. . ;:. PubMed Google Scholar
- Santos-Sierra S. Developments in anticancer vaccination: budding new adjuvants. Biol Chem. 2020;401(4):435-46. . ;:. PubMed Google Scholar
- Seya T, Takeda Y, Matsumoto M. Tumor vaccines with dsRNA adjuvant ARNAX induces antigen-specific tumor shrinkage without cytokinemia. Oncoimmunology. 2016;5(2):e1043506. . ;:. PubMed Google Scholar
- MacDowell KS, Munarriz-Cuezva E, Caso JR, Madrigal JL, Zabala A, Meana JJ, Paliperidone reverts Toll-like receptor 3 signaling pathway activation and cognitive deficits in a maternal immune activation mouse model of schizophrenia. Neuropharmacology. 2017;116:196-207. . ;:. PubMed Google Scholar
- Lebedeva E, Bagaev A, Pichugin A, Chulkina M, Lysenko A, Tutykhina I, The differences in immunoadjuvant mechanisms of TLR3 and TLR4 agonists on the level of antigen-presenting cells during immunization with recombinant adenovirus vector. BMC Immunol. 2018;19(1):26. . ;:. PubMed Google Scholar
- Saxena M, Sabado RL, La Mar M, Mohri H, Salazar AM, Dong H,. Poly-ICLC, a TLR3 Agonist, Induces transient innate immune responses in patients with treated HIV-infection: A randomized double-blinded placebo controlled trial. Front Immunol. 2019;10:725. . ;:. PubMed Google Scholar
- Takeda Y, Kataoka K, Yamagishi J, Ogawa S, Seya T, Matsumoto M. A TLR3-Specific adjuvant relieves innate resistance to PD-L1 blockade without cytokine toxicity in tumor vaccine immunotherapy. Cell Rep. 2017;19(9):1874-87. . ;:. PubMed Google Scholar
- Apostolico JS, Lunardelli VAS, Yamamoto MM, Cunha-Neto E, Boscardin SB, Rosa DS. Poly(I:C) Potentiates T cell immunity to a dendritic cell targeted HIV-multiepitope vaccine. Front Immunol. 2019;10:843. . ;:. PubMed Google Scholar
- Hervas-Stubbs S, Olivier A, Boisgerault F, Thieblemont N, Leclerc C. TLR3 ligand stimulates fully functional memory CD8+ T cells in the absence of CD4+ T-cell help. Blood. 2007;109(12):5318-26. . ;:. PubMed Google Scholar
- Miller SM, Cybulski V, Whitacre M, Bess LS, Livesay MT, Walsh L, Novel Lipidated Imidazoquinoline TLR7/8 Adjuvants Elicit Influenza-Specific Th1 Immune Responses and Protect Against Heterologous H3N2 Influenza Challenge in Mice. Front Immunol. 2020;11:406. . ;:. PubMed Google Scholar
- Kaushik D, Dhingra S, Patil MT, Piplani S, Khanna V, Honda-Okubo Y,. BBIQ, a pure TLR7 agonist, is an effective influenza vaccine adjuvant. Hum Vaccin Immunother. 2020;16(8):1989-96. . ;:. PubMed Google Scholar
- Cervantes JL, Weinerman B, Basole C, Salazar JC. TLR8: the forgotten relative revindicated. Cell Mol Immunol. 2012;9(6):434-8. . ;:. PubMed Google Scholar
- Chen CY, Shih YC, Hung YF, Hsueh YP. Beyond defense: regulation of neuronal morphogenesis and brain functions via Toll-like receptors. J Biomed Sci. 2019;26(1):90. . ;:. PubMed Google Scholar
- Dowling DJ. Recent advances in the discovery and delivery of TLR7/8 agonists as vaccine adjuvants. Immunohorizons. 2018;2(6):185-97. . ;:. PubMed Google Scholar
- Vasilakos JP, Tomai MA. The use of Toll-like receptor 7/8 agonists as vaccine adjuvants. Expert Rev Vaccines. 2013;12(7):809-19. . ;:. PubMed Google Scholar
- Wille-Reece U, Wu CY, Flynn BJ, Kedl RM, Seder RA. Immunization with HIV-1 Gag protein conjugated to a TLR7/8 agonist results in the generation of HIV-1 Gag-specific Th1 and CD8+ T cell responses. J Immunol. 2005;174(12):7676-83. . ;:. PubMed Google Scholar
- Haan JM, Lehar SM, Bevan MJ. CD8(+) but not CD8(-) dendritic cells cross-prime cytotoxic T cells in vivo. J Exp Med. 2000;192(12):1685-96. . ;:. PubMed Google Scholar
- Shortman K, Heath WR. The CD8+ dendritic cell subset. Immunol Rev. 2010;234(1):18-31. . ;:. PubMed Google Scholar
- Schnorrer P, Behrens GM, Wilson NS, Pooley JL, Smith CM, El-Sukkari D, The dominant role of CD8+ dendritic cells in cross-presentation is not dictated by antigen capture. Proc Natl Acad Sci USA. 2006;103(28):10729-34. . ;:. Google Scholar
- Cerovic V, Houston SA, Westlund J, Utriainen L, Davison ES, Scott CL. Lymph-borne CD8alpha+ dendritic cells are uniquely able to cross-prime CD8+ T cells with antigen acquired from intestinal epithelial cells. Mucosal Immunol. 2015;8(1):38-48. . ;:. Google Scholar
- Bonifaz LC, Bonnyay DP, Charalambous A, Darguste DI, Fujii S, Soares H,. In vivo targeting of antigens to maturing dendritic cells via the DEC-205 receptor improves T cell vaccination. J Exp Med. 2004;199(6):815-24. . ;:. Google Scholar
- Idoyaga J, Lubkin A, Fiorese C, Lahoud MH, Caminschi I, Huang Y, et al. Comparable T helper 1 (Th1) and CD8 T-cell immunity by targeting HIV gag p24 to CD8 dendritic cells within antibodies to Langerin, DEC205, and Clec9A. Proc Natl Acad Sci USA. 2011;108(6):2384-9. . ;:. Google Scholar
- Lahoud MH, Ahmet F, Zhang JG, Meuter S, Policheni AN, Kitsoulis S, et al. DEC-205 is a cell surface receptor for CpG oligonucleotides. Proc Natl Acad Sci USA. 2012;109(40):16270-5. . ;:. Google Scholar
- Gong T, Liu L, Jiang W, Zhou R. DAMP-sensing receptors in sterile inflammation and inflammatory diseases. Nat Rev Immunol. 2020;20(2):95-112. . ;:. Google Scholar
- Zindel J, Kubes P. DAMPs, PAMPs, and LAMPs in immunity and sterile inflammation. Annu Rev Pathol. 2020;15:493-518. . ;:. Google Scholar
- De Lorenzo G, Ferrari S, Cervone F, Okun E. Extracellular DAMPs in plants and mammals: Immunity, tissue damage and repair. Trends Immunol. 2018;39(11):937-50. . ;:. Google Scholar
- Gallo PM, Gallucci S. The dendritic cell response to classic, emerging, and homeostatic danger signals. Implications for autoimmunity. Front Immunol. 2013;4:138. . ;:. Google Scholar
- Dumitriu IE, Baruah P, Valentinis B, Voll RE, Herrmann M, Nawroth PP, Release of high mobility group box 1 by dendritic cells controls T cell activation via the receptor for advanced glycation end products. J Immunol. 2005;174(12):7506-15. . ;:. Google Scholar
- Saenz R, Futalan D, Leutenez L, Eekhout F, Fecteau JF, Sundelius S, TLR4-dependent activation of dendritic cells by an HMGB1-derived peptide adjuvant. J Transl Med. 2014;12:211. . ;:. Google Scholar
- Dumitriu IE, Baruah P, Bianchi ME, Manfredi AA, Rovere-Querini P. Requirement of HMGB1 and RAGE for the maturation of human plasmacytoid dendritic cells. Eur J Immunol. 2005;35(7):2184-90. . ;:. Google Scholar
- Apetoh L, Ghiringhelli F, Tesniere A, Criollo A, Ortiz C, Lidereau R, The interaction between HMGB1 and TLR4 dictates the outcome of anticancer chemotherapy and radiotherapy. Immunol Rev. 2007;220:47-59. . ;:. Google Scholar
- Shiratsuchi A, Watanabe I, Takeuchi O, Akira S, Nakanishi Y. Inhibitory effect of Toll-like receptor 4 on fusion between phagosomes and endosomes/lysosomes in macrophages. J Immunol. 2004;172(4):2039-47. . ;:. Google Scholar
- Tang D, Kang R, Livesey KM, Cheh CW, Farkas A, Loughran P, Endogenous HMGB1 regulates autophagy. J Cell Biol. 2010;190(5):881-92. . ;:. Google Scholar
- Tang D, Lotze MT, Kang R, Zeh HJ. Apoptosis promotes early tumorigenesis. Oncogene. 2011;30(16):1851-4. . ;:. Google Scholar
- Tang D, Kang R, Livesey KM, Zeh HJ, Lotze MT. High mobility group box 1 (HMGB1) activates an autophagic response to oxidative stress. Antioxid Redox Signal. 2011;15(8):2185-95. . ;:. Google Scholar
- Nace G, Evankovich J, Eid R, Tsung A. Dendritic cells and damage-associated molecular patterns: endogenous danger signals linking innate and adaptive immunity. 2012;4(1):6-15. . ;:. Google Scholar
- Karacs J, Reithofer M, Kitzmuller C, Kraller M, Schmalz S, Bleichert S, Adjuvants and vaccines used in allergen-specific immunotherapy induce neutrophil extracellular traps. Vaccines (Basel). 2021;9(4). . ;:. Google Scholar
- Stephen J, Scales HE, Benson RA, Erben D, Garside P, Brewer JM. Neutrophil swarming and extracellular trap formation play a significant role in Alum adjuvant activity. NPJ Vaccines. 2017;2:1. . ;:. Google Scholar
- Reithofer M, Karacs J, Strobl J, Kitzmuller C, Polak D, Seif K. Alum triggers infiltration of human neutrophils ex vivo and causes lysosomal destabilization and mitochondrial membrane potential-dependent NET-formation. FASEB J. 2020;34(10):14024-41. . ;:. Google Scholar
- Mitroulis I, Kambas K, Chrysanthopoulou A, Skendros P, Apostolidou E, Kourtzelis I,Neutrophil extracellular trap formation is associated with IL-1beta and autophagy-related signaling in gout. PLoS One. 2011;6(12):e29318. . ;:. Google Scholar
- Hulina A, Grdic Rajkovic M, Jaksic Despot D, Jelic D, Dojder A, Cepelak I. Extracellular Hsp70 induces inflammation and modulates LPS/LTA-stimulated inflammatory response in THP-1 cells. Cell Stress Chaperones. 2018;23(3):373-84. . ;:. Google Scholar
- Somersan S, Larsson M, Fonteneau JF, Basu S, Srivastava P, Bhardwaj N. Primary tumor tissue lysates are enriched in heat shock proteins and induce the maturation of human dendritic cells. J Immunol. 2001;167(9):4844-52. . ;:. Google Scholar
- Srivastava P. Interaction of heat shock proteins with peptides and antigen presenting cells: chaperoning of the innate and adaptive immune responses. 2002;20:395-425. . ;:. Google Scholar
- Salimu J, Spary LK, Al-Taei S, Clayton A, Mason MD, Staffurth J. Cross-Presentation of the oncofetal tumor antigen 5T4 from irradiated prostate cancer cells-A key role for heat-shock protein 70 and receptor CD91. Cancer Immunol Res. 2015;3(6):678-88. . ;:. Google Scholar
- Chen T, Guo J, Han C, Yang M, Cao X. Heat shock protein 70, released from heat-stressed tumor cells, initiates antitumor immunity by inducing tumor cell chemokine production and activating dendritic cells via TLR4 pathway. J Immunol. 2009;182(3):1449-59. . ;:. Google Scholar
- Li W, Lu L, Lu J, Wang X, Yang C, Jin J. cGAS-STING-mediated DNA sensing maintains CD8(+) T cell stemness and promotes antitumor T cell therapy. Sci Transl Med. 2020;12(549). . ;:. Google Scholar
- Wang H, Hu S, Chen X, Shi H, Chen C, Sun L, cGAS is essential for the antitumor effect of immune checkpoint blockade. Proc Natl Acad Sci USA. 2017;114(7):1637-42. . ;:. Google Scholar
- Nicoli F, Paul S, Appay V. Harnessing the induction of CD8(+) T-cell responses through metabolic regulation by pathogen-recognition-receptor triggering in antigen presenting cells. Front Immunol. 2018;9:2372. . ;:. Google Scholar
- Albert ML, Sauter B, Bhardwaj N. Dendritic cells acquire antigen from apoptotic cells and induce class I-restricted CTLs. Nature. 1998;392(6671):86-9. . ;:. PubMed Google Scholar
- Yatim N, Jusforgues-Saklani H, Orozco S, Schulz O, Barreira da Silva R, Reis e Sousa C. RIPK1 and NF-kappaB signaling in dying cells determines cross-priming of CD8(+) T cells. Science. 2015;350(6258):328-34. . ;:. Google Scholar
- Ravindran R, Khan N, Nakaya HI, Li S, Loebbermann J, Maddur MS,. Vaccine activation of the nutrient sensor GCN2 in dendritic cells enhances antigen presentation. Science. 2014;343(6168):313-7. . ;:. Google Scholar
- Kleinnijenhuis J, Quintin J, Preijers F, Joosten LA, Ifrim DC, Saeed S, Bacille Calmette-Guerin induces NOD2-dependent nonspecific protection from reinfection via epigenetic reprogramming of monocytes. Proc Natl Acad Sci USA. 2012;109(43):17537-42. . ;:. Google Scholar
- Arts RJW, Moorlag S, Novakovic B, Li Y, Wang SY, Oosting M,. BCG Vaccination Protects against Experimental Viral Infection in Humans through the Induction of Cytokines Associated with Trained Immunity. Cell Host Microbe. 2018;23(1):89-100 e5. . ;:. Google Scholar
- Netea MG, Joosten LA, Latz E, Mills KH, Natoli G, Stunnenberg HG, Trained immunity: A program of innate immune memory in health and disease. Science. 2016;352(6284):aaf1098. . ;:. Google Scholar
- Netea MG, Latz E, Mills KH, O'Neill LA. Innate immune memory: a paradigm shift in understanding host defense. Nat Immunol. 2015;16(7):675-9. . ;:. Google Scholar
- Kaikkonen MU, Spann NJ, Heinz S, Romanoski CE, Allison KA, Stender JD, Remodeling of the enhancer landscape during macrophage activation is coupled to enhancer transcription. Mol Cell. 2013;51(3):310-25. . ;:. Google Scholar
- Ostuni R, Piccolo V, Barozzi I, Polletti S, Termanini A, Bonifacio S, Latent enhancers activated by stimulation in differentiated cells. Cell. 2013;152(1-2):157-71. . ;:. Google Scholar
- Yoshida K, Maekawa T, Zhu Y, Renard-Guillet C, Chatton B, Inoue K, The transcription factor ATF7 mediates lipopolysaccharide-induced epigenetic changes in macrophages involved in innate immunological memory. Nat Immunol. 2015;16(10):1034-43. . ;:. Google Scholar
- Cheng SC, Quintin J, Cramer RA, Shepardson KM, Saeed S, Kumar V. mTOR- and HIF-1alpha-mediated aerobic glycolysis as metabolic basis for trained immunity. Science. 2014;345(6204):1250684. . ;:. Google Scholar
- Saeed S, Quintin J, Kerstens HH, Rao NA, Aghajanirefah A, Matarese F, Epigenetic programming of monocyte-to-macrophage differentiation and trained innate immunity. Science. 2014;345(6204):1251086. . ;:. Google Scholar
- Khodadadi L, Cheng Q, Radbruch A, Hiepe F. The maintenance of memory plasma cells. Front Immunol. 2019;10:721. . ;:. Google Scholar
- Auner HW, Beham-Schmid C, Dillon N, Sabbattini P. The life span of short-lived plasma cells is partly determined by a block on activation of apoptotic caspases acting in combination with endoplasmic reticulum stress. Blood. 2010;116(18):3445-55. . ;:. Google Scholar
- Crotty S. T Follicular Helper Cell Biology: A decade of discovery and diseases.. Immunity. 2019;50(5):1132-48. . ;:. Google Scholar
- Vinuesa CG, Linterman MA, Yu D, MacLennan IC. Follicular helper T cells. Annu Rev Immunol. 2016;34:335-68. . ;:. Google Scholar
- Crotty S. T follicular helper cell differentiation, function, and roles in disease. Immunity. 2014;41(4):529-42. . ;:. Google Scholar
- Mesin L, Ersching J, Victora GD. Germinal Center B Cell Dynamics. Immunity. 2016;45(3):471-82. . ;:. Google Scholar
- Akkaya M, Kwak K, Pierce SK. B cell memory: building two walls of protection against pathogens. Nat Rev Immunol. 2020;20(4):229-38. . ;:. Google Scholar
- B Cell Development, Activation and effector functions. Primer to the Immune Response . 2014. p. 111-42. . ;:. Google Scholar
- Palm AE, Henry C. Remembrance of things past: Long-term B cell memory after infection and vaccination. Front Immunol. 2019;10:1787. . ;:. Google Scholar
- Perez-Andres M, Paiva B, Nieto WG, Caraux A, Schmitz A, Almeida J, Human peripheral blood B-cell compartments: a crossroad in B-cell traffic. Cytometry B Clin Cytom. 2010;78 Suppl 1:S47-60. . ;:. Google Scholar
- Riedel R, Addo R, Ferreira-Gomes M, Heinz GA, Heinrich F, Kummer J, et al. Discrete populations of isotype-switched memory B lymphocytes are maintained in murine spleen and bone marrow. Nat Commun. 2020;11(1):2570. . ;:. Google Scholar
- Lightman SM, Utley A, Lee KP. Survival of long-lived plasma cells (LLPC): Piecing together the puzzle. Front Immunol. 2019;10:965. . ;:. Google Scholar
- Slifka MK, Antia R, Whitmire JK, Ahmed R. Humoral immunity due to long-lived plasma cells. Immunity. 1998;8(3):363-72. . ;:. Google Scholar
- Kasturi SP, Rasheed MAU, Havenar-Daughton C, Pham M, Legere T, Sher ZJ,. 3M-052, a synthetic TLR-7/8 agonist, induces durable HIV-1 envelope-specific plasma cells and humoral immunity in nonhuman primates. Sci Immunol. 2020;5(48). . ;:. Google Scholar
- Saeed BQ, Al-Shahrabi R, Alhaj SS, Alkokhardi ZM, Adrees AO. Side effects and perceptions following sinopharm COVID-19 vaccination. Int J Infect Dis. 2021. . ;:. Google Scholar
- Ella R, Vadrevu KM, Jogdand H, Prasad S, Reddy S, Sarangi V,Safety and immunogenicity of an inactivated SARS-CoV-2 vaccine, BBV152: a double-blind, randomised, phase 1 trial. The Lancet Infectious Diseases. 2021;21(5):637-46. . ;:. Google Scholar
- Walther W, Stein U. Viral vectors for gene transfer: a review of their use in the treatment of human diseases. Drugs. 2000;60(2):249-71. . ;:. Google Scholar
- Hasanpourghadi M, Novikov M, Ertl HCJ. COVID-19 vaccines based on adenovirus vectors. Trends Biochem Sci. 2021;46(5):429-30. . ;:. Google Scholar
- Anghelina D, Lam E, Falck-Pedersen E. Diminished innate antiviral response to adenovirus vectors in cGAS/STING-deficient mice minimally impacts adaptive immunity. J Virol. 2016;90(13):5915-27. . ;:. Google Scholar
- Ahn J, Barber GN. STING signaling and host defense against microbial infection. Exp Mol Med. 2019;51(12):1-10. . ;:. Google Scholar
- Anwar S, Ul Islam K, Azmi MI, Iqbal J. cGAS-STING-mediated sensing pathways in DNA and RNA virus infections: crosstalk with other sensing pathways. Arch Virol. 2021;166(12):3255-68. . ;:. Google Scholar
- Edwards DK, Jasny E, Yoon H, Horscroft N, Schanen B, Geter T. Adjuvant effects of a sequence-engineered mRNA vaccine: translational profiling demonstrates similar human and murine innate response. J Transl Med. 2017;15(1):1. . ;:. Google Scholar
- Li C, Lee A, Grigoryan L, Arunachalam PS, Scott MKD, Trisal M. Mechanisms of innate and adaptive immunity to the Pfizer-BioNTech BNT162b2 vaccine. Nat Immunol. 2022. . ;:. Google Scholar
- Zhang C, Maruggi G, Shan H, Li J. Advances in mRNA vaccines for infectious diseases. Front Immunol. 2019;10:594. . ;:. Google Scholar
- Teijaro JR, Farber DL. COVID-19 vaccines: modes of immune activation and future challenges. Nat Rev Immunol. 2021;21(4):195-7. . ;:. Google Scholar
- Sahin U, Muik A, Derhovanessian E, Vogler I, Kranz LM, Vormehr M, COVID-19 vaccine BNT162b1 elicits human antibody and TH1 T cell responses. Nature. 2020;586(7830):594-9. . ;:. Google Scholar
- Tarke A, Sidney J, Methot N, Yu ED, Zhang Y, Dan JM, Impact of SARS-CoV-2 variants on the total CD4(+) and CD8(+) T cell reactivity in infected or vaccinated individuals. Cell Rep Med. 2021;2(7):100355. . ;:. Google Scholar
- Reichmuth AM, Oberli MA, Jaklenec A, Langer R, Blankschtein D. mRNA vaccine delivery using lipid nanoparticles. Ther Deliv. 2016;7(5):319-34. . ;:. Google Scholar
- Park KS, Sun X, Aikins ME, Moon JJ. Non-viral COVID-19 vaccine delivery systems. Adv Drug Deliv Rev. 2021;169:137-51. . ;:. PubMed Google Scholar
- Jackson NAC, Kester KE, Casimiro D, Gurunathan S, DeRosa F. The promise of mRNA vaccines: a biotech and industrial perspective. NPJ Vaccines. 2020;5:11. . ;:. PubMed Google Scholar





