
 Open Access
Open Access Abstract
The rubber tree (Hevea brasiliensis) is one of the most valuable members of the genus Hevea because it has high latex production and the latex exploited from the rubber tree is the primary source of natural rubber. Rubber trees are often damaged and exposed to pathogen attacks through wounds during and after tapping. Gene expression studies help us to gain insights into molecular mechanism underlying the responses of this plant species to stress signals such as wounding and pathogenicity. To ensure the reliability of gene expression measurements, quantitative procedures, particularly reverse transcription - quantitative PCR (RT-qPCR), require the normalization of the expression level of a gene of interest (target gene) to that of a stably expressed internal reference gene. Selecting and validating an appropriate reference gene is thus an essential step before conducting RT-qPCR experiments to look for differences in transcriptional expression of the target gene in H. brasiliensis under specific experimental conditions. Here, the transcriptional expression of ubiquitin-conjugating enzyme 2b (HbUBC2b) was measured by RT-qPCR in a set of 12 RNA samples derived from the bark of the rubber tree H. brasiliensis (clone RRIV 209) exposed to mechanical wounding, and stress signal molecules like methyl jasmonate (MeJA). With the aim of estimating PCR efficiency, and thus to include it in further analysis procedures, a standard curve assessment has been developed using HbUBC2b-specific primers and a 5-fold dilution of cDNA as template for qPCR reactions. The amplification efficiency (102,9%), as well as the R2 value (0,9992) are within the recommended range (90-110%). The HbUBC2b gene was shown to be stably expressed across control and treatment groups, and therefore could be used as a reference gene for quantitative gene expression normalization in H. brasiliensis (clone RRIV 209) under mechanical wounding or methyl jasmonate treatment.
MỞ ĐẦU
Cao su Hevea brasiliensis là một loài thân gỗ thuộc họ Đại kích (Euphorbiaceae) và có giá trị kinh tế cao nhất trong chi Hevea . Cây cao su cho năng suất mủ cao. Mủ cao su chứa thành phần chính là cao su tự nhiên – một loại polymer có các đặc tính lý hóa vượt trội 1 . Mủ cao su chứa trong các mạch mủ và được thu hoạch bằng cách sử dụng dao cạo chuyên dụng để tạo vết cắt theo đường xoắn quanh thân, mỗi lần cạo lấy đi một lát vỏ mỏng và lớp cao su nút kín miệng các mạch mủ. Do đó, cây cao su thường xuyên bị tổn thương và phơi nhiễm với sự tấn công của mầm bệnh thông qua vết thương trong và giữa các lần cạo lấy mủ. Để đáp ứng với các tổn thương cơ học và sự phơi nhiễm với các loài côn trùng và nấm gây bệnh qua vết thương, thực vật nói chung và cây cao su nói riêng thường tổng hợp jasmonic acid (JA) và các dẫn xuất của JA (gọi chung là jasmonate) như một tín hiệu tự vệ. Đáng chú ý, các gene trong con đường truyền tín hiệu jasmonate được kích hoạt rất mạnh trong các tế bào chứa mủ và đặc biệt là trong đáp ứng với stress do quá trình cạo lấy mủ cao su 2 . Ngoài ra, một loạt các gene liên quan đến con đường sinh tổng hợp JA và con đường sinh tổng hợp terpenoid đã được ghi nhận có biểu hiện cao ở các cây khỏe mạnh, nhưng biểu hiện thấp ở những cây bị bệnh “khô miệng cạo” (tapping panel dryness) và có năng suất cho mủ giảm 3 .
Việc phân tích các tác động của tổn thương cơ học và tín hiệu stress JA đến sự biểu hiện của các gene liên quan đến quá trình sinh tổng hợp mủ cao su và đáp ứng stress của cây cao su có thể góp phần làm sáng tỏ cơ chế phân tử của những quá trình này. Northern blotting, cDNA microarray và RT-qPCR là những kỹ thuật phân tích biểu hiện gene thông dụng 4 , 5 , 6 . Những năm gần đây, với sự ra đời của các công nghệ giải trình tự thông lượng cao, kỹ thuật RNA-seq trở thành một công cụ mới trong nghiên cứu biểu hiện gene nói chung và trong các phân tích ảnh hướng của stress đến sự biểu hiện phiên mã của các gene quan tâm ở cây cao su H. brasiliensis nói riêng 7 , 8 , 9 . Tuy vậy, RNA-seq là một kỹ thuật có chi phí cao, lại cần nhiều thời gian để thiết kế, thực hiện và phân tích dữ liệu. Trong khi đó, phương pháp định lượng RT-qPCR hiện vẫn đang được sử dụng phổ biến vì dễ thực hiện, chi phí thấp, có độ nhạy và độ chính xác cao 5 , 10 , 11 , 12 , 13 . Độ chính xác của RT-qPCR phụ thuộc vào nhiều yếu tố như chất lượng RNA tách chiết, hiệu suất tổng hợp cDNA từ mRNA thông qua phiên mã ngược, tính đặc hiệu của cặp mồi, hiệu suất của quá trình nhân bản… 11 , 14 , 15 . Để giảm thiểu ảnh hưởng của những yếu tố vừa nêu đến tính chính xác của kết quả định lượng, quy trình định lượng RT-qPCR luôn bao gồm bước chuẩn hóa dữ liệu biểu hiện của các gene đích với dữ liệu biểu hiện của các gene tham chiếu (reference gene) 11 , 16 .
Trên lý thuyết, gene tham chiếu được định nghĩa là gene có mô hình biểu hiện ổn định trong các mô, các kiểu gene và các điều kiện thí nghiệm khác nhau. Một số gene “giữ nhà” (housekeeping genes) như actin ( ACT ), elongation factor 1-α ( eEF-1α ) ở sinh vật nhân chuẩn, 18S rRNA , glyceraldehyde-3-phosphate dehydrogenase ( GAPDH ) và polyubiquitin ( UBQ )… đã được sử dụng rộng rãi làm gene tham chiếu trong các phân tích RT-qPCR 17 . Tuy nhiên, ngày càng có thêm nhiều dữ liệu cho thấy mô hình biểu hiện phiên mã của các gene “giữ nhà” có thể biến động trong nhiều trường hợp 18 , 19 , 20 , 21 và do đó, mỗi gene “giữ nhà” chỉ có thể được sử dụng làm gene tham chiếu ở những điều kiện mà gene đó có biểu hiện ổn định 22 , 23 , 24 . Điều này nhấn mạnh tầm quan trọng của việc xác nhận tính ổn định của gene tham chiếu được chọn cho mỗi hệ thống sinh học trước khi thực hiện khảo sát sự biểu hiện gene quan tâm trên hệ thống đó 25 , 26 , 27 .
Sự phát triển của kỹ thuật RNA-seq thông lượng cao tạo cơ hội cho việc sàng lọc sơ bộ các gene có biểu hiện phiên mã ổn định để sử dụng làm gene tham chiếu. Kết quả đánh giá ban đầu về độ ổn định biểu hiện của 23 gene tham chiếu tiềm năng bằng các thuật toán geNorm và NormFinder trên dữ liệu từ 18 mẫu đại diện cho giai đoạn đầu, giữa và cuối của mạch mủ ở hai giống cao su có thời gian cho mủ khác nhau cho thấy hai gene thành viên của họ gene ubiquitin-conjugating enzyme (UBC), gồm UBC2a và UBC2b, có biểu hiện ổn định nhất 28 . UBC2a và UBC2b cũng biểu hiện ổn định ở các mô/tế bào khác nhau, bao gồm cả trong các tế bào chứa mủ (laticifer) và mô vỏ của cây cao su. Ngoài ra, UBC2b là một trong hai gene ổn định nhất khi so sánh biểu hiện của gene giữa các cá thể cây và giữa các cây cao su mang kiểu gene khác nhau 29 . Gene này cũng có độ ổn định cao trong nhiều điều kiện thí nghiệm khác, ví dụ như khi cây được xử lý hormone hay trong suốt quá trình tái tạo mủ giữa hai lần thu hoạch mủ 29 , 30 . Trong báo cáo này, phần vỏ của thân cây cao su RRIV 209 được tạo vết cắt cơ học sâu vào vùng tế bào sinh mủ và methyl jasmonate được sử dụng để mô phỏng tín hiệu stress nhằm đánh giá tính ổn định trong sự biểu hiện của gene HbUBC2b ở mô vỏ khi cây cao su chịu tổn thương cơ học hoặc khi được xử lý với MeJA ngoại sinh.
VẬT LIỆU VÀ PHƯƠNG PHÁP
Vật liệu nghiên cứu
Giống cao su ( Hevea brasiliensis ) RRIV 209 được trồng tại Viện Nghiên cứu Cao su Việt Nam ( xã Lai Hưng, huyện Bàu Bàng, tỉnh Bình Dương).
Phương pháp xử lý tạo vết thương cơ học ở phần vỏ cây cao su Hevea brasiliensis
Các cây cao su RRIV 209 được trồng trong cùng một điều kiện tại Viện Nghiên cứu Cao su Việt Nam. Cụ thể, 18 cây cao su 3 tháng tuổi có kích thước đồng đều được lựa chọn cho hai nghiệm thức: xử lý tạo vết thương (Wounding test – WT) và đối chứng (Wounding control – WC). Mỗi nghiệm thức có 3 tổ hợp, với mỗi tổ hợp bao gồm 3 cây cao su. Các cây ở lô xử lý được tạo 5 vết cắt sâu vào vùng tế bào sinh mủ ở phần vỏ tại vị trí giữa hai tầng lá 12 . Các vết cắt song song và cách nhau 0,25 cm ( Figure 1 ). Các cây ở lô đối chứng không bị gây tổn thương. Sau 4 giờ xử lý, mô vỏ thân của các cây xử lý và đối chứng được thu nhận và bảo quản trong nitrogen lỏng.
Figure 1 . Xử lý tạo vết thương cơ học trên cây cao su H. brasiliensis (giống RRIV 209)

Phương pháp xử lý cây cao su Hevea brasiliensis với MeJA
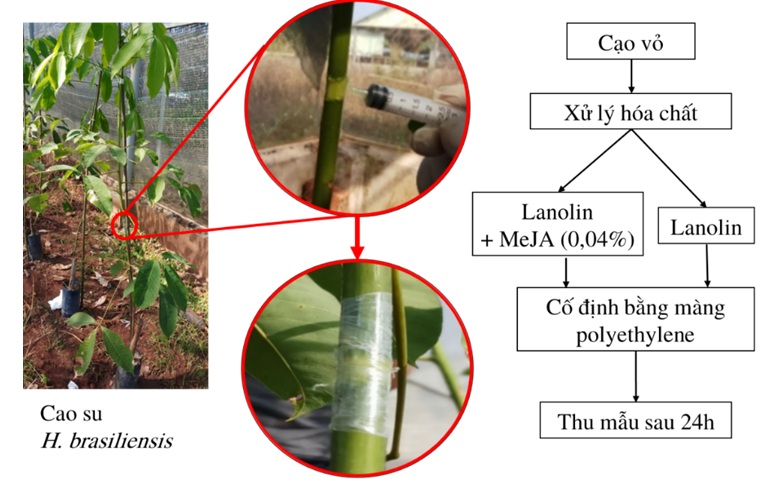
Tương tự, 18 cây cao su RRIV 209 3 tháng tuổi có kích thước đồng đều được lựa chọn cho hai nghiệm thức: xử lý MeJA (Jasmonate test – JT) và đối chứng (Jasmonate control – JC). Mỗi nghiệm thức gồm 3 tổ hợp, với mỗi tổ hợp bao gồm 3 cây cao su. Cụ thể, tiến hành cạo bỏ lớp biểu bì và một phần tầng sinh vỏ vòng quanh thân cây tại vị trí giữa hai tầng lá, chiều cao của vòng cạo khoảng 1 cm. Sau đó, 0,1 mL hỗn hợp gồm lanolin và MeJA 0,04% (w/w) được thoa lên vết cạo của các cây của lô xử lý. Vị trí xử lý được cố định bằng cách bọc một lớp màng polyethylene ( Figure 2 ). Lô đối chứng được thực hiện tương tự nhưng phần thân đã cạo được thoa hỗn hợp chỉ chứa lanolin 31 . Sau 24 giờ xử lý, mô vỏ của đoạn thân dài khoảng 2,5 cm cách phía trên và phía dưới vùng xử lý 0,5 cm được thu nhận và trữ trong nitrogen lỏng.
Tách chiết RNA tổng số và tổng hợp cDNA
Mẫu vỏ thân cao su RRIV 209 từ mỗi tổ hợp được nghiền thành dạng bột trong nitrogen lỏng. Khoảng 50 mg bột mô vỏ được sử dụng để tách chiết RNA tổng số bằng kít EZ-10 spin column Plant RNA Mini-preps (Bio Basic, Canada). Các dụng cụ và hóa chất liên quan đến thao tác trên RNA được xử lý với H 2 O 2 3% và DEPC 1‰ để loại bỏ hoàn toàn RNase. Quy trình loại bỏ DNA bộ gene được thực hiện bằng kít RapidOut DNA Removal (Thermo Fisher Scientific, USA). Tính toàn vẹn của RNA được kiểm tra bằng phương pháp điện di trên gel agarose 1%. Chất lượng RNA được đánh giá dựa trên tỷ lệ của sự hấp thụ tại bước sóng 260 và 280 nm (A 260 /A 280 ). Hàm lượng RNA trong các mẫu được xác định bằng phương pháp đo mật độ quang thông qua máy quang phổ Nano 7415 (Jenway, UK). Quá trình tổng hợp cDNA được thực hiện theo hướng dẫn của kít Reverse transcriptase Aid First Strand (Thermo Fisher Scientific, USA), sử dụng 1 µg RNA cho phản ứng có thể tích 20 µL. cDNA được dùng làm khuôn cho các phản ứng qPCR.
Thiết kế cặp mồi nhân bản đặc hiệu đoạn amplicon từ bản sao của gene HbUBC2b
Mồi xuôi HbUBC2b-qF và mồi ngược HbUBC2b-qR được thiết kế bằng phần mềm OligoAnalyzer 3.1 32 với các thông số được khuyến nghị như sau: mồi có chiều dài 18-30 bp, hàm lượng GC trong khoảng 30-80%, nhiệt độ nóng chảy (melting temperature, T m ) của mồi trong khoảng 55-65 o C. Nhiệt độ nóng chảy của mồi xuôi và mồi ngược không chênh nhau quá 5 o C. Mồi không chứa cấu trúc kẹp tóc (hairpin loop) và không hình thành dimer. Sản phẩm nhân bản (amplicon) có kích thước 75-150 bp 33 . Ít nhất một trong hai mồi bắt cặp bổ sung với trình tự vắt ngang vị trí nối giữa hai exon kề nhau, để mồi chỉ bắt cặp đặc hiệu với cDNA mà không thể liên kết với DNA bộ gen. Tính đặc hiệu của cặp mồi với trình tự gene tham chiếu được dự đoán bằng công cụ primer-BLAST của NCBI 34 .
Khảo sát nhiệt độ gắn mồi trong phản ứng real-time PCR
Thí nghiệm khảo sát nhiệt độ gắn mồi được thực hiện ở 5 điểm nhiệt độ 54,8 o C, 58,2 o C, 62,5 o C, 64,7 o C và 68,3 o C. Phản ứng real-time PCR (tổng thể tích 10 μL) được thực hiện trên hệ thống real-time PCR LightCycler® 96 Instrument (Roche, Germany) với thành phần như sau: 6,2 μL nước đã qua xử lý DEPC; 1 μL đệm phản ứng h-Taq 10X; 0,2 μL MgCl 2 10 mM; 0,2 μL dNTP 10 mM; 0,4 μL mồi xuôi HbUBC2b-qF 10 mM; 0,4 μL mồi ngược HbUBC2b-qR 10 mM; 0,5 μL EvaGreen® Dye 20X; 0,1 μL SolGent™ h-Taq DNA polymerase 2,5 U/μl; 1 μL cDNA. Chu trình nhiệt của phản ứng real-time PCR được thiết lập theo hướng dẫn của kít h-Taq DNA Polymerase (SolGent™, Korea): 95 o C/ 15 phút, 40 chu kỳ của 95 o C (20 giây), T a o C (40 giây) và 72 o C (6 giây), trong đó T a (annealing temperature) là nhiệt độ gắn mồi. Để kiểm tra tín hiệu huỳnh quang tạo ra từ phản ứng qPCR là do sản phẩm nhân bản đặc hiệu, tiến hành phân tích đường cong nóng chảy (melting curve) sau khi phản ứng kết thúc với chương trình nhiệt độ như sau: 95 o C/10 giây; 65 o C/60 giây và tăng dần nhiệt độ đến 97 o C, mỗi lần tăng 0,2 o C, giữ lại trong 1 giây và đọc tín hiệu. Nhiệt độ gắn mồi được chọn để thực hiện các khảo sát tiếp theo là nhiệt độ tại đó chỉ xuất hiện một đỉnh tín hiệu huỳnh quang trong khoảng 75-90 o C trên đường cong nóng chảy và phản ứng có giá trị chu kỳ ngưỡng (C t ) thấp nhất.
Phân tích trình tự đoạn gene được nhân bản
Sản phẩm nhân bản của phản ứng real-time PCR sử dụng cặp mồi HbUBC2b-qF và HbUBC2b-qR được điện di trên gel agarose 1%, được tinh sạch và tạo dòng vào vector pJET1.2-blunt (Thermo Fisher Scientific, USA). Plasmid mang trình tự amplicon được giải trình tự với mồi pJET1.2-F.
Khảo sát hiệu suất nhân bản
Hiệu suất nhân bản với cặp mồi đặc hiệu HbUBC2b-qF và HbUBC2b-qR tại nhiệt độ gắn mồi đã khảo sát được đánh giá thông qua đường chuẩn biểu diễn mối tương quan giữa chu kỳ ngưỡng (C t ) với số lượng bản sao trong các mẫu cDNA đã được pha loãng theo hệ số 5. Hệ số tương quan (R 2 ) của đường chuẩn phản ánh độ chính xác và độ lặp lại của thí nghiệm. Hiệu suất nhân bản (E%) được tính toán theo công thức: E% = (n -1/slope -1) × 100, với n là hệ số pha loãng cDNA và slope là độ dốc của đường chuẩn 35 , 36 . Phản ứng RT-qPCR cho kết quả định lượng tin cậy khi hệ số tương quan R 2 ≥ 0,985 và hiệu suất nhân bản (E%) trong khoảng 90-110% 35 , 37 .
Khảo sát sự biểu hiện của gene HbUBC2b ở điều kiện xử lý gây tổn thương và xử lý với MeJA
Mức biểu hiện phiên mã của gene HbUBC2b được khảo sát bằng qPCR trên 12 mẫu cDNA tổng hợp từ RNA tổng số thu nhận từ mô vỏ của cây cao su RRIV 209 được xử lý gây tổn thương hoặc xử lý với methyl jasmonate (MeJA) và của các cây đối chứng tương ứng. Giá trị chu kỳ ngưỡng C t trong hai nghiệm thức đối chứng và xử lý ở mỗi thí nghiệm được ghi nhận và so sánh bằng phân tích thống kê.
Xử lý thống kê
Giá trị chu kỳ ngưỡng Ct tb của các phản ứng real-time PCR có nhiệt độ gắn mồi khác nhau được so sánh bằng phương pháp ANOVA một yếu tố và phương pháp Tukey HSD. Mức biểu hiện của gene HbUBC2b ở mẫu xử lý (được thể hiện qua giá trị chu kỳ ngưỡng C t ) được so sánh với ở mẫu đối chứng bằng phương pháp Student’s t-test (two-tail, unpaired, equal variance).
KẾT QUẢ VÀ THẢO LUẬN
T hiết kế cặp mồi nhân bản đặc hiệu đoạn amplicon từ bản sao của gene tham chiếu HbUBC2b
Các thông số của mồi và vị trí bắt cặp của mồi xuôi HbUBC2b-qF và mồi ngược HbUBC2b-qR trên trình tự bản sao HbUBC2b được trình bày ở Table 1 và Figure 3 . Kích thước dự đoán của sản phẩm nhân bản ( HbUBC2b _ amplicon ) là 93 bp . Kết quả kiểm tra tính đặc hiệu của cặp mồi với trình tự đích bằng chương trình Primer-Blast của NCBI được thể hiện ở Figure 4 . Theo đó, cặp mồi HbUBC2b-qF và HbUBC2b-qR không chỉ bắt cặp với trình tự mục tiêu là HbUBC2b (Genbank accession number: XM_021809251) mà còn bắt cặp với ba trình tự khác. Tuy nhiên, do nucleotide ở tận cùng đầu 3’ của mồi xuôi HbUBC2b-qF không bổ sung với vị trí tương ứng trên ba trình tự đó nên phản ứng real-time PCR với cặp mồi HbUBC2b-qF và HbUBC2b-qR được dự đoán là chỉ nhân bản đặc hiệu trình tự đích HbUBC2b_amplicon ( Figure 4 ).
Figure 3 . Vị trí của mồi xuôi HbUBC2b-qF và mồi ngược HbUBC2b-qR trên trình tự bản sao HbUBC2b . Vùng trình tự được tô nền và không được tô nền xen kẽ nhau tương ứng với các exon kề nhau

Figure 4 . Kết quả kiểm tra độ đặc hiệu của cặp mồi HbUBC2b-qF và HbUBC2b-qR bằng chương trình Primer-Blast

Tách chiết RNA tổng số
Kết quả điện di các mẫu RNA tách chiết từ các tổ hợp mẫu vỏ sau khi đã xử lý loại DNA bộ gene được thể hiện ở Figure 5 . Theo đó, các mẫu RNA đều có chất lượng tốt với các vạch 18S và 28S hiện diện rõ nét.
Figure 5 . Kết quả điện di mẫu RNA tổng số tách chiết từ mô vỏ cây cao su H. brasiliensis (giống RRIV 209) trong thí nghiệm xử lý gây tổn thương (A) và thí nghiệm xử lý MeJA (B)

Khảo sát nhiệt độ gắn mồi trong phản ứng qPCR
Để xác định nhiệt độ lai tối ưu cho sự bắt cặp của cặp mồi HbUBC2b-qF và HbUBC2b-qR với trình tự đích, phản ứng real-time PCR được thực hiện tại 5 điểm nhiệt độ 54,8 o C, 58,2 o C, 62,5 o C, 64,7 o C và 68,3 o C. Kết quả cho thấy các đường cong nóng chảy ứng với các điểm nhiệt độ khảo sát trên đều chỉ có một đỉnh tín hiệu huỳnh quang duy nhất trong khoảng nhiệt độ 78-84 o C ( Figure 6 ), được cho là tương ứng với sản phẩm nhân bản dự đoán HbUBC2b_amplicon . So sánh giá trị chu kỳ ngưỡng Ct tb giữa các phản ứng real-time PCR có nhiệt độ gắn mồi khác nhau bằng phương pháp ANOVA một yếu tố và phương pháp Tukey HSD cho thấy với nhiệt độ gắn mồi 62,5 o C hoặc 64,7 o C ( Table 2 , Figure 6 ) phản ứng real-time PCR cho giá trị Ct tb nhỏ hơn so với giá trị Ct tb của các phản ứng sử dụng các nhiệt độ gắn mồi còn lại (P = 2,88.10 -7 ). Trong các thí nghiệm tiếp theo, chọn 64,7 o C làm nhiệt độ gắn mồi.
Figure 6 . Đường cong nóng chảy của sản phẩm nhân bản trong các phản ứng real-time PCR sử dụng các nhiệt độ gắn mồi (T a ) khác nhau

Phân tích trình tự của đoạn gene được nhân bản (amplicon)
Sản phẩm nhân bản của phản ứng real-time PCR sử dụng cặp mồi HbUBC2b-qF và HbUBC2b-qR được tinh sạch và giải mã trình tự. Đầu tiên, sản phẩm nhân bản được điện di trên gel agarose 1% ( Figure 7 ). Theo đó, ở giếng 3 của bản gel có xuất hiện một vạch nằm dưới vạch có kích thước 250 bp của thang chuẩn DNA (Thermo Fisher Scientific, USA), phù hợp với kích thước dự đoán 93 bp của sản phẩm nhân bản đặc hiệu ( HbUBC2b _ amplicon ). Bên cạnh đó, ở đối chứng âm (giếng 1) (sử dụng nước cất thay cho cDNA) không có xuất hiện vạch có kích thước tương ứng. Sản phẩm nhân bản ở giếng 3 Figure 7 được tinh sạch từ gel agarose và được tạo dòng vào vector pJET1.2-blunt. Plasmid tái tổ hợp mang trình tự amplicon được giải trình tự với mồi pJET1.2-F. Kết quả giải trình tự ( Figure 9 ) cho thấy đoạn amplicon dòng hóa giống 100% với trình tự của sản phẩm nhân bản dự đoán ( HbUBC2b_amplicon ) và do đó đây là sản phẩm nhân bản duy nhất và đặc hiệu của phản ứng .
Figure 7 . Kết quả điện di sản phẩm real-time PCR sử dụng cặp mồi HbUBC2b-qF và HbUBC2b-qR trên gel agarose 1%. Giếng 1: Đối chứng âm không bổ sung cDNA; Giếng 2: Thang chuẩn GeneRuler 1kb DNA Ladder (Thermo Fisher Scientific); Giếng 3: Sản phẩm nhân bản sử dụng khuôn cDNA chuẩn bị từ mô vỏ của H. brasiliensis

Hiệu suất nhân bản trình tự amplicon từ bản sao của gene HbUBC2b bằng qPCR
Hiệu suất nhân bản HbUBC2b_amplicon được đánh giá thông qua đường chuẩn biểu diễn mối tương quan giữa số lượng bản sao trong các mẫu cDNA đã được pha loãng theo hệ số 5 và giá trị chu kỳ ngưỡng (C t ) trung bình tương ứng ( Figure 8 ). Đồ thị đường chuẩn có hệ số tương quan R 2 = 0,9992 và có phương trình y= -2,2747 x + 23,177, trong đó độ dốc ( slope ) = -2,2747. Hiệu suất nhân bản của phản ứng qPCR được tính toán theo công thức: E% = (5 -1/slope -1) × 100 = 102,9%. Như vậy, hiệu suất nhân bản nằm trong phạm vi khuyến nghị (90-110%) đảm bảo độ tin cậy của kết quả định lượng 35 , 36 , 37 .
Figure 8 . Đồ thị đường chuẩn biểu diễn mối tương quan giữa chu kỳ ngưỡng (C t ) và nồng độ cDNA khi thực hiện phản ứng qPCR nhân bản HbUBC2b _amplicon

Biểu hiện phiên mã của gene HbUBC2b khi cây cao su có vết thương cơ học hoặc được xử lý MeJA
Trước khi sử dụng HbUBC2b làm gene tham chiếu cho các thí nghiệm phân tích biểu hiện phiên mã của các gene quan tâm khi cây cao su bị gây tổn thương cơ học hoặc bị xử lý MeJA, gene HbUBC2b cần được xác nhận có biểu hiện ổn định ở điều kiện này hay không. Giá trị C t của các phản ứng qPCR sử dụng cặp mồi HbUBC2b-qF và HbUBC2b-qR trên các mẫu cDNA từ thí nghiệm xử lý gây tổn thương và xử lý MeJA (mỗi thí nghiệm gồm 2 nghiệm thức: xử lý và đối chứng) được trình bày ở Table 3 và Table 4 .
Kết quả phân tích thống kê so sánh giá trị chu kỳ ngưỡng C t trong hai nghiệm thức ở mỗi thí nghiệm bằng phương pháp t -test (two-tail, unpaired, equal variance) cho thấy không có sự khác biệt có ý nghĩa thống kê giữa mẫu xử lý và mẫu đối chứng (P = 0,3191 và P = 0,6916). Điều này chứng tỏ mức biểu hiện phiên mã của gene HbUBC2b không thay đổi khi cây cao su có tổn thương cơ học hoặc được xử lý MeJA. Như vậy, HbUBC2b là gene tham chiếu phù hợp cho thí nghiệm khảo sát sự biểu hiện của các gene quan tâm ở hai điều kiện xử lý trên. Tương tự, các nghiên cứu sàng lọc các gene tham chiếu dựa trên so sánh độ ổn định biểu hiện của chúng giữa các cá thể cây, các kiểu gene, các loại mô / tế bào, các phân đoạn khác nhau của mạch mủ và trong một số điều kiện thí nghiệm khác đã công bố cũng ghi nhận rằng gene mã hóa ubiquitin-conjugating enzyme 2b ( HbUBC2b ) là một trong các gene có độ ổn định cao nhất ở H. brasiliensis 28 , 29 , 30 .
KẾT LUẬN
Ubiquitin-conjugating enzyme 2b ( HbUBC2b) là gene tham chiếu phù hợp cho các phân tích biểu hiện gene bằng phương pháp RT-qPCR ở vỏ cây cao su RRIV 209 khi cây đáp ứng với tổn thương cơ học hoặc chịu tác động của MeJA. Phản ứng qPCR có thành phần phản ứng gồm 6,2 μL nước đã qua xử lý DEPC; 1 μL đệm phản ứng h-Taq 10X; 0,2 μL MgCl 2 10 mM; 0,2 μL dNTP 10 mM; 0,4 μL mồi xuôi HbUBC2b-qF 10 mM; 0,4 μL mồi ngược HbUBC2b-qR 10 mM; 0,5 μL EvaGreen® Dye 20X; 0,1 μL SolGent™ h-Taq DNA polymerase 2,5 U/μL; và 1 μL cDNA, được thực hiện ở chu kỳ nhiệt sau: 95 o C/ 15 phút, 40 chu kỳ của 95 o C (20 giây), 64,7 o C (40 giây) và 72 o C (6 giây) đã cho phép nhân bản đặc hiệu trình tự HbUBC2b_amplicon có kích thước 93 bp với hiệu suất nhân bản (102,9%) trong khoảng khuyến nghị 90-110%.
PHỤ LỤC
Figure 9 . Kết quả giải trình tự plasmid pJET1.2- amplicon với mồi pJET1.2-F. Sản phẩm nhân bản HbUBC2b_amplicon được dòng hóa vào vector tương ứng với đoạn trình tự từ nucleotide 36 đến 129

DANH MỤC CÁC TỪ VIẾT TẮT
DEPC: diethyl pyrocarbonate
dNTP: deoxynucleotide triphosphate
HSD: honestly significant difference
JA: jasmonic acid
MeJA: methyl jasmonate
RRIV: Rubber Research Institute of Vietnam
RT-qPCR: reverse transcriptase – quantitative polymerase chain reaction
UBC: ubiquitin-conjugating enzyme
XUNG ĐỘT LỢI ÍCH
Các tác giả không có xung đột lợi ích nào liên quan đến các kết quả đã công bố.
ĐÓNG GÓP CỦA CÁC TÁC GIẢ
Trần Thị Diễm Hương lên kế hoạch nghiên cứu, thực hiện các thí nghiệm, thu thập và xử lý dữ liệu. Nguyễn Thành Long thực hiện các thí nghiệm, thu thập và xử lý dữ liệu. Huỳnh Bích Trâm thực hiện các thí nghiệm và hỗ trợ xử lý dữ liệu. Trần Thanh chuẩn bị vật liệu nghiên cứu và thảo luận góp ý phương pháp nghiên cứu. Nguyễn Thị Hồng Thương định hướng và lên kế hoạch nghiên cứu, hướng dẫn nghiên cứu, viết và hoàn thiện bản thảo.
LỜI CẢM ƠN
Nghiên cứu được tài trợ bởi Đại học Quốc gia Thành phố Hồ Chí Minh (ĐHQG-HCM) trong khuôn khổ Đề tài mã số B2019-18-2. Nhóm nghiên cứu chân thành cảm ơn Huỳnh Nguyễn Minh Nghĩa và TS. Nguyễn Xuân Dũng - Phòng CNSH Thực vật, Trung tâm Công nghệ Sinh học Thành phố Hồ Chí Minh đã hỗ trợ quá trình thực hiện thí nghiệm qPCR, và TS. Nguyễn Tiến Dũng – Bộ môn Toán ứng dụng, Trường Đại học Bách Khoa – ĐHQG HCM đã hỗ trợ phân tích thống kê.
References
- Lau N-S, Makita Y, Kawashima M, Taylor TD, Kondo S, Othman AS, et al. The rubber tree genome shows expansion of gene family associated with rubber biosynthesis. Sci Rep [Internet]. 2016 Sep 24;6(1):28594. . ;:. Google Scholar
- Pirrello J, Leclercq J, Dessailly F, Rio M, Piyatrakul P, Kuswanhadi K, et al. Transcriptional and post-transcriptional regulation of the jasmonate signalling pathway in response to abiotic and harvesting stress in Hevea brasiliensis. BMC Plant Biol [Internet]. 2014 Dec 2;14(1):341. . ;:. Google Scholar
- Liu J-P, Xia Z-Q, Tian X-Y, Li Y-J. Transcriptome sequencing and analysis of rubber tree (Hevea brasiliensis Muell.) to discover putative genes associated with tapping panel dryness (TPD). BMC Genomics [Internet]. 2015 Dec 21;16(1):398. . ;:. Google Scholar
- Roth CM. Quantifying gene expression. Curr Issues Mol Biol [Internet]. 2002 Jul;4(3):93-100. . ;:. Google Scholar
- VanGuilder HD, Vrana KE, Freeman WM. Twenty-five years of quantitative PCR for gene expression analysis. Biotechniques [Internet]. 2008 Apr;44(5):619-26. . ;:. PubMed Google Scholar
- Schena M, Shalon D, Davis RW, Brown PO. Quantitative Monitoring of Gene Expression Patterns with a Complementary DNA Microarray. Science (80- ) [Internet]. 1995 Oct 20;270(5235):467-70. . ;:. PubMed Google Scholar
- Mohamed Sathik MB, Luke LP, Rajamani A, Kuruvilla L, Sumesh K V, Thomas M. De novo transcriptome analysis of abiotic stress-responsive transcripts of Hevea brasiliensis. Mol Breed [Internet]. 2018 Mar 22;38(3):32. . ;:. Google Scholar
- Campos Mantello C, Boatwright L, da Silva CC, Scaloppi EJ, de Souza Goncalves P, Barbazuk WB, et al. Deep expression analysis reveals distinct cold-response strategies in rubber tree (Hevea brasiliensis). BMC Genomics [Internet]. 2019 Dec 4;20(1):455. . ;:. PubMed Google Scholar
- Wilhelm BT, Landry J-R. RNA-Seq-quantitative measurement of expression through massively parallel RNA-sequencing. Methods [Internet]. 2009 Jul;48(3):249-57. . ;:. PubMed Google Scholar
- Derveaux S, Vandesompele J, Hellemans J. How to do successful gene expression analysis using real-time PCR. Methods [Internet]. 2010 Apr;50(4):227-30. . ;:. PubMed Google Scholar
- Huggett J, Dheda K, Bustin S, Zumla A. Real-time RT-PCR normalisation; strategies and considerations. Genes Immun [Internet]. 2005 Jun 7;6(4):279-84. . ;:. PubMed Google Scholar
- Putranto R-A, Duan C, Kuswanhadi, Chaidamsari T, Rio M, Piyatrakul P, et al. Ethylene Response Factors Are Controlled by Multiple Harvesting Stresses in Hevea brasiliensis. Fu B, editor. PLoS One [Internet]. 2015 Apr 23;10(4):e0123618. . ;:. PubMed Google Scholar
- Wu C, Lan L, Li Y, Nie Z, Zeng R. The relationship between latex metabolism gene expression with rubber yield and related traits in Hevea brasiliensis. BMC Genomics. . 2018;:. PubMed Google Scholar
- Nolan T, Hands RE, Bustin SA. Quantification of mRNA using real-time RT-PCR. Nat Protoc [Internet]. 2006 Aug 9;1(3):1559-82. . ;:. PubMed Google Scholar
- Pfaffl MW. Quantification strategies in real-time PCR. In: A-Z of quantitative PCR. . 2004;:. Google Scholar
- Livak KJ, Schmittgen TD. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods [Internet]. 2001 Dec;25(4):402-8. . ;:. PubMed Google Scholar
- Schmidt GW, Delaney SK. Stable internal reference genes for normalization of real-time RT-PCR in tobacco (Nicotiana tabacum) during development and abiotic stress. Mol Genet Genomics [Internet]. 2010 Mar 23;283(3):233-41. . ;:. PubMed Google Scholar
- Dheda K, Huggett JF, Bustin SA, Johnson MA, Rook G, Zumla A. Validation of housekeeping genes for normalizing RNA expression in real-time PCR. Biotechniques [Internet]. 2004 Jul;37(1):112-9. . ;:. PubMed Google Scholar
- Suzuki T, Higgins PJ, Crawford DR. Control Selection for RNA Quantitation. Biotechniques [Internet]. 2000 Aug;29(2):332-7. . ;:. PubMed Google Scholar
- Bustin S. Absolute quantification of mRNA using real-time reverse transcription polymerase chain reaction assays. J Mol Endocrinol [Internet]. 2000 Oct 1;25(2):169-93. . ;:. PubMed Google Scholar
- Basa B, Solti Á, Sárvári É, Tamás L. Housekeeping gene selection in poplar plants under Cd-stress: comparative study for real-time PCR normalisation. Funct Plant Biol [Internet]. 2009;36(12):1079. . ;:. PubMed Google Scholar
- Radonić A, Thulke S, Mackay IM, Landt O, Siegert W, Nitsche A. Guideline to reference gene selection for quantitative real-time PCR. Biochem Biophys Res Commun [Internet]. 2004 Jan;313(4):856-62. . ;:. PubMed Google Scholar
- Huis R, Hawkins S, Neutelings G. Selection of reference genes for quantitative gene expression normalization in flax (Linum usitatissimum L.). BMC Plant Biol [Internet]. 2010;10(1):71. . ;:. PubMed Google Scholar
- Thellin O, Zorzi W, Lakaye B, De Borman B, Coumans B, Hennen G, et al. Housekeeping genes as internal standards: use and limits. J Biotechnol [Internet]. 1999 Oct;75(2-3):291-5. . ;:. Google Scholar
- Bustin SA, Beaulieu J-F, Huggett J, Jaggi R, Kibenge FSB, Olsvik PA, et al. MIQE précis: Practical implementation of minimum standard guidelines for fluorescence-based quantitative real-time PCR experiments. BMC Mol Biol [Internet]. 2010;11(1):74. . ;:. PubMed Google Scholar
- Remans T, Keunen E, Bex GJ, Smeets K, Vangronsveld J, Cuypers A. Reliable Gene Expression Analysis by Reverse Transcription-Quantitative PCR: Reporting and Minimizing the Uncertainty in Data Accuracy. Plant Cell [Internet]. 2014 Oct;26(10):3829-37. . ;:. PubMed Google Scholar
- Chapman JR, Waldenström J. With Reference to Reference Genes: A Systematic Review of Endogenous Controls in Gene Expression Studies. PLoS One [Internet]. 2015 Nov 10;10(11):e0141853. . ;:. PubMed Google Scholar
- Chao J, Yang S, Chen Y, Tian W-M. Evaluation of Reference Genes for Quantitative Real-Time PCR Analysis of the Gene Expression in Laticifers on the Basis of Latex Flow in Rubber Tree (Hevea brasiliensis Muell. Arg.). Front Plant Sci [Internet]. 2016 Jul 29;7. . ;:. Google Scholar
- Li H, Qin Y, Xiao X, Tang C. Screening of valid reference genes for real-time RT-PCR data normalization in Hevea brasiliensis and expression validation of a sucrose transporter gene HbSUT3. Plant Sci [Internet]. 2011 Aug;181(2):132-9. . ;:. PubMed Google Scholar
- Long X, He B, Gao X, Qin Y, Yang J, Fang Y, et al. Validation of reference genes for quantitative real-time PCR during latex regeneration in rubber tree. Gene [Internet]. 2015 Jun;563(2):190-5. . ;:. PubMed Google Scholar
- Liu J-P, Hu J, Liu Y-H, Yang C-P, Zhuang Y-F, Guo X-L, et al. Transcriptome analysis of Hevea brasiliensis in response to exogenous methyl jasmonate provides novel insights into regulation of jasmonate-elicited rubber biosynthesis. Physiol Mol Biol Plants [Internet]. 2018 May 13;24(3):349-58. . ;:. PubMed Google Scholar
- Owczarzy R, Tataurov A V., Wu Y, Manthey JA, McQuisten KA, Almabrazi HG, et al. IDT SciTools: a suite for analysis and design of nucleic acid oligomers. Nucleic Acids Res. 2008. . ;:. PubMed Google Scholar
- Livak KJ. Allelic discrimination using fluorogenic probes and the 5′ nuclease assay. Genet Anal Biomol Eng [Internet]. 1999 Feb;14(5-6):143-9. . ;:. Google Scholar
- Ye J, Coulouris G, Zaretskaya I, Cutcutache I, Rozen S, Madden TL. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinformatics [Internet]. 2012 Dec 18;13(1):134. . ;:. PubMed Google Scholar
- Svec D, Tichopad A, Novosadova V, Pfaffl MW, Kubista M. How good is a PCR efficiency estimate: Recommendations for precise and robust qPCR efficiency assessments. Biomol Detect Quantif [Internet]. 2015 Mar;3:9-16. . ;:. PubMed Google Scholar
- Töwe S, Kleineidam K, Schloter M. Differences in amplification efficiency of standard curves in quantitative real-time PCR assays and consequences for gene quantification in environmental samples. J Microbiol Methods [Internet]. 2010 Sep;82(3):338-41. . ;:. PubMed Google Scholar
- Rogers-Broadway KR, Karteris E. Amplification efficiency and thermal stability of qPCR instrumentation: Current landscape and future perspectives. Exp Ther Med [Internet]. 2015 Oct;10(4):1261-4. . ;:. PubMed Google Scholar





