
 Open Access
Open Access Abstract
Saltwater intrusion in the Mekong Delt, an urgent problem, affected the growth of fruit trees in general and Citrus latifolia (Yu. Tanaka) Tanaka in particular. Sodium chloride in saline soils cause the water stress on plant roots leading to the leaf senescence and death of plant. Therefore, in this study, the salinity tolerance of the studied plant was investigated under conditions of additional sodium chloride in soil at various concentrations (from 0 to 16 g/L) to determine the salinity tolerance threshold of the plants, and then the roots were water irrigated and the leaves were sprayed by KNO3 solution at concentrations from 0 to 10 g /L to improve the salinity tolerance. The results showed that after 2 weeks of treatment of sodium chloride 4‒16 g/L, the increased saline caused the osmotic stress, increased the defoliation but reduced the leaf renewal, the fresh weight, the chlorophyll index, the Citrus latifolia growth, and decreased the aperture size and the photosynthetic intensity over time. The salinity stress promoted the ligninization in the rhizosphere parenchyma cells, reduced the root renewal and elongation. In addition, treatments for the plant recovery were also carried out by irrigating at the roots and spraying KNO3 10 g/L solution on the leaves. After 3 and 6 weeks, irrigation treatments could only recover plants from sodium chloride at 4‒8 g/L, indicating by the new leaf formation, the maintenance of the chlorophyll content in leaves and the decrease of the proline content. Sodium chloride solutions of 12 and 16 g/L caused a complete death of plants which could not be recovered after 6 weeks of watering. The treatment of sodium chloride combining with KNO3 10 g/L spray helped the aperture opening and increasing the proline accumulation in leaves and roots. In addition, role of endogenous plant growth regulators was also discussed to clarify physiological changes in saline stress conditions.
MỞ ĐẦU
Chanh không hạt Citrus latifolia (Yu. Tanaka) Tanaka là cây ăn quả có chứa ascorbic acid, sucrose và một số khoáng như calcium, potassium và manganese..., được trồng với diện tích hàng nghìn hecta tại đồng bằng sông Cửu Long 1 . Tuy nhiên gần đây, đồng bằng sông Cửu Long đang chịu ảnh hưởng của hiện tượng xâm nhập mặn, nước mặn lấn sâu vào đất liền khoảng 30–75 km, gây hại đến việc canh tác cây chanh không hạt 2 . Khi gặp stress mặn, cây bị xáo trộn cân bằng nước, sự hấp thu của rễ bị cản, lá sẽ bị hóa vàng. Tùy vào loài thực vật mà khả năng đáp ứng với stress mặn sẽ khác nhau như tăng sự lignin hóa vách tế bào làm tăng độ bền cơ học hay tăng hàm lượng proline góp phần làm tăng áp suất thẩm thấu của tế bào 3 , 4 , 5 , 6 , 7 . Trong giới hạn bài báo này, chúng tôi khảo sát khả năng chịu mặn của cây chanh không hạt bằng cách xử lý tưới dung dịch NaCl ở các nồng độ khác nhau. Sau đó xử lý tưới nước rửa mặn ở rễ và phun dung dịch KNO 3 lên lá nhằm góp phần tìm ra giải pháp hạn chế tác hại của sự nhiễm mặn lên sự sinh trưởng của cây chanh không hạt.
VẬT LIỆU VÀ PHƯƠNG PHÁP
Vật liệu
Cây chanh không hạt dạng nhánh chiết (8 tuần tuổi tính từ lúc cắt khỏi cây mẹ, tại huyện Đức Hòa, tỉnh Long An) có chiều cao 40 ± 5 cm, mang 1–3 nhánh phụ ( Figure 1 ), được trồng trong vườn thực nghiệm (Thủ Đức, Thành phố Hồ Chí Minh).
Với các thí nghiệm trong vườn thực nghiệm, cây chanh không hạt được trồng trong chậu nhựa (thể tích 1,5 lít), chứa 750 ± 20 g giá thể bao gồm xơ dừa : đất trồng : trấu (tỉ lệ 1 : 2 : 1 theo trọng lượng) đã được rửa chát và phơi khô (độ ẩm khoảng 40 %). Đáy chậu được lót bằng lớp nilong mỏng có đục lỗ để giá thể thoát nước
Phương pháp
Khảo sát ảnh hưởng của các độ mặn khác nhau lên sự tăng trưởng của cây chanh không hạt chiết nhánh
Cây chanh không hạt được tưới dung dịch NaCl ở các nồng độ: 0, 4, 8, 12 và 16 g/L, bằng cách rót đều lên bề mặt giá thể, với thể tích 250 mL/cây/lần tưới, vào các thời điểm 8 giờ và 16 giờ mỗi ngày (tháng 04/2017). Điều kiện trong vườn thực nghiệm: nhiệt độ ngày 33 ± 2 o C, ánh sáng 32,2 ± 15,1 klux, độ ẩm 54 ± 7 %.
Các chỉ tiêu như tỉ lệ lá rụng, chỉ số diệp lục tố (Chlorophyll Content Index), độ mở khẩu và cường độ quang hợp của lá, hàm lượng proline, trọng lượng tươi, hoạt tính chất điều hòa tăng trưởng thực vật (ĐHTTTV) nội sinh được ghi nhận.
Khảo sát ảnh hưởng của nước lên khả năng phục hồi của cây chanh không hạt chiết nhánh
Cây chanh không hạt sau ba tuần được xử lý NaCl với nồng độ từ 0‒16 g/L tiếp tục được xử lý phục hồi bằng cách tưới nước ở rễ, thể tích 250 mL/cây/lần tưới, vào các thời điểm 8 giờ và 16 giờ mỗi ngày (tháng 05/2017). Thí nghiệm được theo dõi trong khoảng thời gian 3 và 6 tuần sau khi bắt đầu tưới nước phục hồi, các kết quả như số lá tạo mới, chỉ số diệp lục tố, hàm lượng proline được ghi nhận.
Khảo sát ảnh hưởng KNO3 lên sự tăng trưởng và phục hồi của cây chanh không hạt khi nhiễm mặn
Cây chanh không hạt được xử lý riêng lẻ với NaCl 0, 8 và 16 g/L bằng cách tưới dung dịch lên giá thể, vào các thời điểm 8 giờ và 16 giờ mỗi ngày. Ngoài ra, xử lý phun dung dịch KNO 3 10 g/L (phun trực tiếp đến khi ướt đều lá) được kết hợp cùng với xử lý tưới dung dịch NaCl 0, 8 và 16 g/L, thể tích 75 mL/cây/lần phun (tại thời điểm 7 giờ sáng), 7 lần phun trên một thí nghiệm, mỗi lần cách nhau 3 ngày. Các nghiệm thức, bao gồm:
● (i) Xử lý tưới NaCl, 0 g/L
●((ii) Xử lý tưới NaCl, 8 g/L
● (iii) Xử lý tưới NaCl, 16 g/L
● (iv) Xử lý tưới NaCl, 8 g/L và phun KNO 3 , 10 g/L
●(v) Xử lý tưới NaCl, 16 g/L và phun KNO 3 , 10 g/L
Thí nghiệm được theo dõi 2 tuần, các kết quả như chỉ số diệp lục tố, độ mở khẩu và cường độ quang hợp của lá, hàm lượng proline được ghi nhận.
Quan sát cấu trúc giải phẫu
Phẫu thức ngang của lá và rễ cây chanh không hạt được quan sát trực tiếp trong nước cất và sau khi nhuộm hai màu đỏ carmin–xanh iod bằng kính hiển vi quang học.
Xác định độ mở khẩu
Các mẫu lá (tại vị trí cách gốc 30 cm) được thoa lớp keo mỏng để cố định trạng thái các khí khẩu. Độ mở khẩu là khoảng cách chính giữa 2 vách trong của tế bào khí khẩu, được ghi nhận bằng phần mềm LAS V4.0.
Đo cường độ quang hợp
Cường độ quang hợp của lá (µmol O 2 /m 2 /giây) được xác định bằng máy đo trao đổi khí oxygen (Hansatech, Anh) ở điều kiện ánh sáng 30 ± 2 klux và nhiệt độ buồng đo duy trì 26 ± 1 o C nhờ hệ thống giải nhiệt.
Xác định chỉ số diệp lục tố (Chlorophyll Content Index)
Chỉ số diệp lục tố (CCI) được đo trực tiếp bằng máy Chlorophyll Content Meter- CCM 200 plus (hãng Opti Sciences, Mỹ), dựa vào nguyên tắc phổ hấp thu của diệp lục tố tại bước sóng 653 nm và 931 nm.
Xác định hàm lượng proline trong mẫu
Dịch trích proline thu được từ lá và rễ được cho phản ứng với hỗn hợp gồm ninhydrin 1 % (w/v), acetic acid 60 % (v/v) và ethanol 20 % (v/v) với tỉ lệ 1 : 2 (theo thể tích), trong điều kiện nhiệt độ 95 o C (20 phút). Dung dịch được để nguội trong điều kiện phòng và đo mật độ quang ở bước sóng 520 nm. Ghi nhận các giá trị đo và tính hàm lượng proline trong mẫu dựa vào đường chuẩn.
Đo hoạt tính chất điều hòa tăng trưởng thực vật (ĐHTTTV) nội sinh
Các hợp chất ĐHTTTV nội sinh trong mẫu lá và rễ được ly trích và phân tích trên bản mỏng silica gel, trong hệ dung môi isopropanol: ammonium hydroxyde: H 2 O với tỉ lệ 10:1:1 (theo thể tích). Hoạt tính của IAA và ABA được đo bằng sinh trắc nghiệm với diệp tiêu của lúa Oryza sativa (L.). Hoạt tính của GA được đo bằng sinh trắc nghiệm với cây mầm xà lách Lactuca sativa (L.).
Bố trí thí nghiệm và xử lý thống kê
Các thí nghiệm được bố trí ngẫu nhiên với 3 lần lặp lại, mỗi nghiệm thức gồm 9 mẫu. Số liệu được xử lý phân tích thống kê bằng phép thử Duncan và T-Test (one way ANOVA) nhờ phần mềm SPSS (Statistical Package for Social Sciences), phiên bản 22.0 dùng cho Windows. Sự khác biệt có ý nghĩa ở mức p £0,05 của các giá trị được biểu hiện bằng các mẫu tự khác nhau kèm theo sau.
KẾT QUẢ VÀ THẢO LUẬN
Ảnh hưởng của các mức độ mặn khác nhau lên sự tăng trưởng của cây chanh không hạt chiết nhánh.
Sự thay đổi trong cấu trúc giải phẫu của rễ
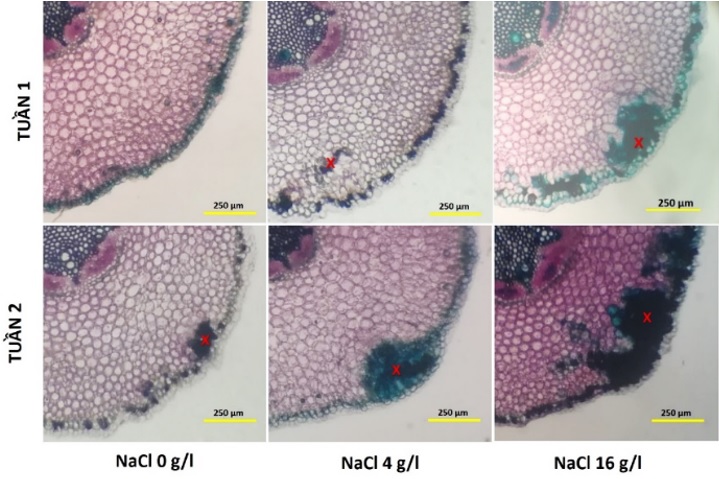
Cắt ngang qua vùng lông hút của rễ cho thấy bên trong tầng lông hút là một lớp tế bào có vách dày và hóa suberin, nhu mô vỏ vách còn cenluloz. Sau một tuần xử lý tưới NaCl với nồng độ từ 0‒16 g/L thì các tế bào nhu mô vỏ gần tầng lông hút có vách hóa lignin tăng dần theo nồng độ NaCl. Đặc biệt, sự hóa ligin của tế bào nhu mô vỏ tăng mạnh sau 2 tuần xử lý ở nồng độ NaCl 16 g/L ( Figure 2 ). Trong điều kiện stress muối, nồng độ NaCl cao gây ra stress thẩm thấu, thường làm xáo trộn cân bằng nước của thực vật và cản sự hấp thu nước của rễ 8 . Do đó đáp ứng của rễ nhằm tránh tổn hại do stress muối trong thí nghiệm này là biểu hiện tăng sự lignin hóa làm tăng độ bền cơ học để chống chịu tốt với stress giúp cây thích ứng với điều kiện sống 9 , 3 , 7 .

Figure 2 . Phẫu thức ngang rễ của cây chanh không hạt sau 1 và 2 tuần xử lý NaCl ở các nồng độ 0, 4 và 16 g/L có sự tẩm lignin ở các tế bào nhu mô vỏ (x)
Sự hóa vàng và sự rụng của lá theo thời gian
Trên cây bị xử lý NaCl 4 g/L sau 2 tuần, ở vùng phiến lá gần cuống bắt đầu bằng sự xuất hiện vùng màu xanh nhạt và sau đó chuyển sang màu vàng. Sự hóa vàng này tăng theo thời gian và nồng độ xử lý NaCl. Khi tăng nồng độ NaCl 4 đến 16 g/ L thì sự vàng lá có xu hướng lan rộng theo phần mép về phía ngọn lá và sau đó lan theo gân phụ vào vùng gân chính. Sau 3 tuần xử lý NaCl, ngoài sự hóa vàng thì lá bắt đầu có sự hóa nâu, với biểu hiện là vùng hóa vàng ở phiến lá gần gân chính xuất hiện những vết màu nâu và khi tăng nồng độ NaCl 4 đến 16 g/L thì vết hóa nâu xuất hiện nhiều hơn ở 2 mép lá, mép lá bị khô, co lại và rụng ( Figure 3 ).
Phẫu thức ngang vùng lá hóa vàng cho thấy lớp lục mô rào gần biểu bì trên và lớp lục mô khuyết gần biểu bì dưới của lá có lục lạp nhạt màu và có xu hướng chuyển sang vàng. Bên cạnh đó, độ dày của lá giảm so với trước khi xử lý, do sự thu hẹp các khoảng gian bào ở lục mô khuyết ( Figure 4 A và Figure 4 B). Lá bị hóa hoàng là triệu chứng phổ biến nhất thể hiện khi cây bị stress mặn 10 và ở chanh không hạt cũng không ngoại lệ.Trong thí nghiệm xử lý stress muối NaCl trực tiếp xâm nhập vào bộ rễ, sự thay đổi cấu trúc ở rễ như đã trình bày ở trên giúp cây chống chịu với stress thẩm thấu, làm hạn chế sự hấp thu nước vào bên trong mạch mộc và dẫn lên lá. Trong điều kiện thiếu nước do stress muối gây ra, lá bắt đầu có biểu hiện giảm độ dày thông qua sự thu hẹp các các khoảng gian bào ở vùng tế bào lục mô khuyết ( Figure 4 B). Sự hóa vàng và hóa nâu của lá trong trường hợp này là do sự nhạt màu xanh lục ở lá, lá bắt đầu tổn hại do thiếu nước, từ đó thành lập vùng rụng làm tỉ lệ rụng lá tăng cao khi xử lý NaCl ở nồng độ 4-16 g/L.
Figure 3 . Lá trưởng thành cây chanh không hạt tại vị trí cách gốc 25 cm, sau 2 và 3 tuần xử lý NaCl ở các nồng độ khác nhau: 0 (A), 4 (B), 8 (C), 12 (D) và 16 (E) g/L

Figure 4 . Phẫu thức ngang của lá cách gốc 25 cm bị hoàng hóa khi xử lý NaCl trong vườn thực nghiệm (A) và chi tiết vùng mô gần biểu bì trên với lục mô rào và lục mô khuyết có lục lạp nhạt màu, nhu mô khuyết mất màu xanh (B)

Sau 2 tuần xử lý NaCl, tỉ lệ rụng lá của cây ở nồng độ NaCl 4 và 8 g/L không có sự khác biệt với đối chứng (NaCl 0 g/L), nhưng tỉ lệ này bắt đầu tăng ở nồng độ NaCl 12 g/L và 16 g/L. Sau 3 tuần xử lý, tỉ lệ rụng lá tăng từ nồng độ NaCl 8 g/L trở lên. Ngoài ra, kết quả cho thấy tỉ lệ rụng lá ở tuần thứ 3 tăng so với tuần thứ 2 và tăng mạnh ở cây bị xử lý NaCl 12 g/L (tăng 34,54 %) và 16 g/L (tăng 32,38 %) ( Figure 5 ). Thực tế cho thấy cây ở nồng độ NaCl 0 g/L và 4 g/L có sự tạo mới của lá ở ngọn, làm tăng tổng số lá trên cây ở tuần thứ 2 sau xử lý. Theo thời gian, sự rụng lá ở cây được xử lý NaCl xảy ra tuần tự, bắt đầu từ phần gần gốc trở lên ( Figure 6 ).
Figure 5 . Ảnh hưởng của NaCl ở các nồng độ khác nhau lên tỉ lệ rụng lá (%) trên cây chanh không hạt sau 2 và 3 tuần xử lý.

Figure 6 . Cây chiết nhánh chanh không hạt sau 2 tuần xử lý NaCl ở các nồng độ khác nhau: 0 (A), 4 (B), 8 (C), 12 (D) và 16 (E) g/l.

Trong thí nghiệm xử lý NaCl với các nồng độ khác nhau, ở nồng độ NaCl 0 g/L vẫn có sự rụng lá do hiện tượng lão suy của các lá trưởng thành. Thay đổi cấu trúc tế bào có ý nghĩa trong sự lão suy lá là sự suy thoái lục lạp. Sự hóa vàng hay hóa nâu ở lá cho thấy sự phá hủy hay lục lạp bị tách rời trong lão suy lá có ý nghĩa quan trọng với sự huy động lại chất dinh dưỡng để tạo lại cơ quan mới 8 . Do đó, sự tăng tỉ lệ rụng lá cũng tương ứng với xu hướng tăng diện tích hóa vàng và hóa nâu trên lá, điều này làm giảm chỉ số diệp lục tố ở lá khi cây bị xử lý NaCl từ 4‒16 g/L ( Figure 3 ). Sau 3 tuần xử lý tưới NaCl với các nồng độ tăng dần từ 0‒16 g/L, trọng lượng tươi của lá và rễ cũng giảm theo ( Figure 7 ). Thực tế cho thấy, rễ cây chanh không hạt ở nồng độ NaCl 0 và 4 g/L có sự tạo mới của rễ và rễ kéo dài ở nồng độ 0 g/L. Khi tăng nồng độ NaCl thì sinh khối của rễ giảm ( Figure 8 ).
Figure 7 . Ảnh hưởng của NaCl theo nồng độ lên trọng lượng tươi (g) của lá và rễ trên cây chanh không hạt sau 3 tuần xử lý
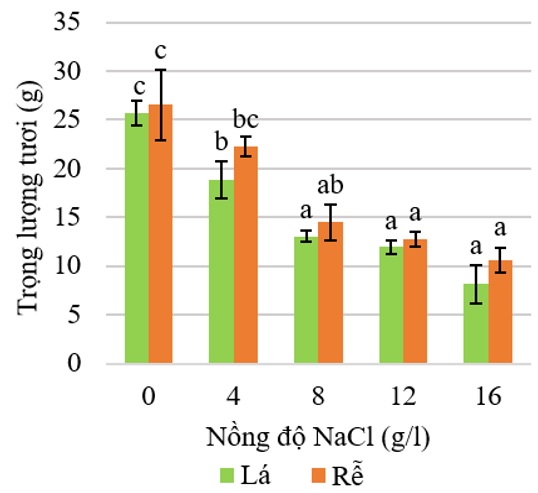
Figure 8 . Rễ cây chanh không hạt sau 3 tuần xử lý NaCl ở các nồng độ khác nhau: 0 (A), 4 (B), 8 (C), 12 (D) và 16 (E) g/L

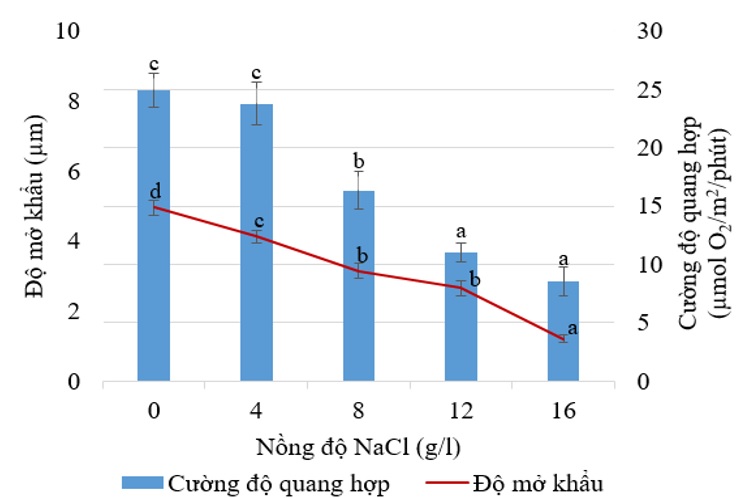
Tương tự như chỉ số diệp lục tố ở lá, cây chanh không hạt sau 2 tuần xử lý NaCl, có độ mở khẩu của lá bắt đầu giảm khi xử lý tưới NaCl nồng độ 4 g/L trở lên ( Figure 9 ). Cùng với sự giảm chỉ số diệp lục tố ở lá, giảm cường độ quang hợp đã làm cây giảm tăng trưởng hoặc tăng trưởng chậm khi stress muối kéo dài. Khả năng quang hợp giảm do stress muối cũng được ghi nhận ở cây đậu Phaseolus vulgaris (L.) 11 . Sau 2 tuần xử lý NaCl, cường độ quang hợp đầu giảm ở nồng độ NaCl 8-16 g/L ( Figure 10 ). Stress muối gây giảm sự hấp thụ nước do sự hóa lignin ở rễ và sự giảm độ mở khí khẩu ở lá ( Figure 11 B), từ đó ngăn chặn dòng nước mang các ion độc như Cl - vào tế bào gây tổn hại cho các mô, đồng thời điều đó cũng làm giảm sự hấp thu CO 2 8 . Sự thiếu CO 2 do stress muối gây ra làm giảm sự tiêu thụ NADPH bởi chu trình Calvin dẫn đến ferredoxin bị khử quá mức trong chuỗi chuyển điện tử quang hợp và điện tử sẽ được chuyển từ PSI đến phân tử oxygen, từ đó hình thành gốc tự do O 2 - . Sự sản xuất dư thừa các gốc tự do gây ra sự oxy hóa các thành phần trong tế bào như lipid, protein, acid nucleic và gây phá hủy bộ máy quang hợp 12 .
Ở những cây có xử lý NaCl thì hàm lượng proline tích lũy ở lá cao hơn hẳn. Ngoài ra, mặc dù với lượng thấp hơn so với lá, nhưng hàm lượng proline tích lũy của rễ cũng tăng cao. Ở cả lá và rễ, hàm lượng proline đều đạt mức cao nhất trên cây ở nồng độ NaCl 12 g/L và cao hơn so với cây ở các nồng độ khác ( Figure 11 A). Dưới điều kiện stress muối, cây có xu hướng tăng tổng hợp và tích lũy proline trong tế bào chất 8 . Trong thí nghiệm này, tăng hàm lượng proline góp phần làm tăng áp suất thẩm thấu của tế bào, giúp tế bào tăng khả năng hấp thu nước. Phân tử proline trong tế bào chất hoạt động để duy trì áp suất thẩm thấu cao trong tế bào nhằm cải thiện khả năng hấp thu nước vào tế bào chất 4 .
Figure 9 . Ảnh hưởng của NaCl ở các nồng độ khác nhau lên chỉ số diệp lục tố của lá (CCI) trên cây chanh không hạt sau 2 và 3 tuần xử lý.

Figure 10 . Ảnh hưởng của NaCl ở các nồng độ khác nhau lên cường độ quang hợp của lá (µmol O 2 /m 2 /phút) trên cây chanh không hạt sau 2 tuần xử lý.
Cây chanh không hạt được xử lý tưới với dung dịch NaCl nồng độ từ 0‒16 g/L sau 2 tuần, có sự thay đổi hoạt tính các chất điều hòa tăng trưởng thực vật nội sinh ở lá và rễ. Các chất điều hòa tăng trưởng thực vật nội sinh đóng vai trò quyết định khả năng chống chịu của thực vật đối với nhiều loại stress bao gồm stress muối. Hoạt tính IAA tăng cao khi tăng nồng độ NaCl từ 0‒16 g/L ( Figure 11 B), có thể liên quan đến sự giảm tăng trưởng của cây 13 . Kết quả này góp phần giải thích cho hiện tượng ức chế sự tạo mới và sự kéo dài rễ ( Figure 8 ). GA giúp kéo dài tế bào thực vật. Hoạt tính Gibberellin của lá và rễ cây chanh không hạt đều giảm sau 2 tuần xử lý NaCl 0‒16 g/L ( Figure 11 C). Hoạt tính ABA của lá và rễ đều tăng cao khi tăng nồng độ NaCl xử lý 4‒16 g/L ( Figure 11 D), kết quả này tương tự như kết quả nghiên cứu ghi nhận trên cây lúa 14 và đậu nành 15 . Nhiều nghiên cứu cho thấy ABA đóng vai trò như chất truyền tín hiệu nội bào, cảm ứng sự biểu hiện các gien đáp ứng với stress 13 , 16 . Hoạt tính ABA tăng liên quan đến sự đóng khí khẩu, tăng tích lũy proline và protein 17 .
Figure 11 . Ảnh hưởng của NaCl ở các nồng độ khác nhau lên hàm lượng proline của lá (màu xanh) và rễ (màu đỏ) trên cây chanh không hạt sau 2 tuần xử lý ( Figure 11 A), hoạt tính của các chất IAA ( Figure 11 B), GA ( Figure 11 C) và ABA ( Figure 11 D) nội sinh ở lá và rễ cây chanh không hạt sau 2 tuần xử lý NaCl ở các nồng độ khác nhau.

Ảnh hưởng của tưới nước rửa mặn lên khả năng phục hồi của cây chanh không hạt chiết nhánh
Cây chanh không hạt sau 3 tuần tưới NaCl được xử lý phục hồi bằng cách tưới nước rửa mặn trong giá thể cho thấy các cây ở nồng độ NaCl 0, 4 và 8 g/L có sự hình thành lá mới theo thời gian. Các cây được xử lý ở nồng độ NaCl 12 và 16 g/L không có khả năng tạo mới lá. Sau 6 tuần, khả năng tạo lá mới của cây được xử lý ở nồng độ NaCl 4 và 8 g/L thấp hơn so với NaCl 0 g/L, còn ở nồng độ NaCl 12 và 16 g/L cây bị rụng lá hoàn toàn ( Figure 12 ). Việc tưới nước giúp rửa trôi hàm lượng muối NaCl trong đất, làm giảm sự ảnh hưởng trực tiếp của nồng độ muối cao và điều chỉnh áp suất thẩm thấu. Tuy nhiên ở tuần thứ 3, các cây được xử lý ở nồng độ NaCl 12 và 16 g/L đã rụng toàn bộ lá trên cây, không tạo lá mới, sinh khối rễ giảm mạnh và bị hư gần như toàn bộ. Trong khi ở nồng độ NaCl 4 và 8 g/L vẫn có lá tạo mới, mặc dù thấp hơn so với nồng độ NaCl 0 g/L. Điều này có thể được giải thích thông qua sự tăng nồng độ muối thì đã làm hư các nhu mô ở rễ và giảm số lượng rễ trước đó ( Figure 13 ), từ đó làm giảm khả năng hấp thụ nước, dẫn đến cây chết sau 6 tuần phục hồi ở xử lý với NaCl nồng độ cao (12 và 16 g/L). Sau 3 tuần phục hồi, ngoại trừ các cây được xử lý ở nồng độ NaCl 12 g/L vẫn tiếp tục có sự giảm chỉ số diệp lục tố, còn ở các cây khác vẫn tiếp tục duy trì và không có sự khác biệt so với thời điểm xử lý NaCl 3 tuần (tương ứng tuần 0 khi bắt đầu xử lý phục hồi). Bên cạnh đó, chỉ số diệp lục tố không khác biệt theo thời gian phục hồi, nhưng trên các cây có xử lý NaCl 4‒16 g/L thì vẫn thấp hơn so với cây không xử lý (NaCl 0 g/L) ( Figure 14 ).
Figure 12 . Cây chanh không hạt sau 6 tuần phục hồi có sự tạo mới lá của cây ở các nồng độ NaCl 0 (A), 4 (B) và 8 (C) g/L; các cây ở nồng độ NaCl 12 (D) và 16 (E) g/L lá rụng hoàn toàn.

Figure 13 . Rễ cây chanh không hạt sau 6 tuần phục hồi có sự thay đổi sinh khối rễ trên các cây đã xử lý NaCl nồng độ 0 (A), 4 (B), 8 (C), 12 (D) và 16 (E) g/L lá rụng hoàn toàn sau 6 tuần phục hồi

Figure 14 . Chỉ số diệp lục tố (CCI) của lá cây chanh sau 3 và 6 tuần xử lý tưới nước rửa mặn.

Figure 15 . Hàm lượng proline tích lũy (mg.l -1 /mg trọng lượng tươi) ở lá và rễ trên cây chanh không hạt sau 6 tuần phục hồi.

Sau khi tưới nước rửa mặn ở rễ, các cây ở nồng độ NaCl 4 và 8 g/L có sự tạo mới của lá và duy trì chỉ số diệp lục tố, điều này có thể giải thích là do nước vẫn có thể tiếp tục được dẫn lên theo mạch do sự hư hoại các nhu mô vùng vỏ và phần chóp rễ còn non, do số lượng rễ còn lại ít ( Figure 13 ) và hoạt động sinh lý giảm do stress thẩm thấu gây ra, làm cho sự tăng trưởng trong quá trình phục hồi của cây cũng thấp hơn so với đối chứng (NaCl 0 g/L). Sau khi xử lý tưới nước rửa mặn, chỉ số diệp lục tố của lá đối với cây đã xử lý NaCl vẫn thấp hơn so với cây đối chứng là do hiện tượng các lá trưởng thành vẫn tiếp tục rụng và gần như rụng hoàn toàn với cây đã chịu nồng độ muối cao trước đó (nồng độ NaCl 12 và 16 g/L). Hàm lượng proline ở các cây sau khi phục hồi 6 tuần giảm xuống thấp hơn với các cây đang được xử lý NaCl. Tuy nhiên, hàm lượng proline ở lá và rễ của cây ở nồng độ NaCl 8 g/L thì lại tăng so với trước khi phục hồi. Sau 3 tuần phục hồi, hàm lượng proline tích lũy trên lá có sự tương đương giữa cây được xử lý ở nồng độ NaCl 0 và 4 g/L. Còn đối với rễ, sự tích lũy này khác biệt giữa các cây được xử lý ở nồng độ NaCl 0, 4 và 8 g/L. Bên cạnh đó, hàm lượng proline ở rễ vẫn thấp hơn so với lá ( Figure 15 ).
Ảnh hưởng của việc xử lý tưới NaCl riêng lẻ hoặc kết hợp với phun KNO3 lên sự tăng trưởng của cây chanh không hạt
Xử lý riêng lẻ tưới NaCl hoặc kết hợp phun KNO 3 đều làm giảm chỉ số diệp lục tố (CCI) của lá sau 2 tuần. Trong đó, cây khi xử lý ở nồng độ NaCl 16 g/L có chỉ số diệp lục tố thấp nhất và khi xử lý NaCl 8 hoặc 16 g/L kết hợp phun KNO 3 10 g/L cho thấy chỉ số diệp lục tố cao hơn so với xử lý riêng lẻ NaCl 8 và 16 g/l ( Table 1 ).
Hàm lượng proline (mg. L -1 /mg trọng lượng tươi) tích lũy cao hơn ở lá trưởng thành so với rễ. Tăng nồng độ khi xử lý riêng lẻ NaCl thì làm tăng hàm lượng proline tích lũy ở rễ và cao nhất ở nồng độ 16 g/L. Xử lý NaCl 8 và 16 g/L kết hợp phun KNO 3 làm tăng cao proline tích lũy ở cả rễ và lá ( Table 2 ).
Trong các tế bào thực vật, ion potasssium vẫn giữ trạng thái cân bằng ion, và đóng vai trò của một chất đa lượng. K + giúp cân bằng ion (điều hòa thế thẩm thấu), cử động khí khẩu 8 . K + được cung cấp bằng việc phun trực tiếp KNO 3 (10 g/L) qua lá, giúp lá nhanh chóng hấp thu K + , sự hấp thu ion khoáng này cùng với sự tăng hàm lượng proline tích lũy ( Table 2 ) dẫn đến sự tăng áp suất thẩm thấu của tế bào của lá và rễ, độ mở khí khẩu ở lá được phun KNO 3 cũng tăng, giúp cho khả năng thoát hơi nước ở lá và dòng nước được hấp thu từ rễ lên lá cũng tăng theo. Khí khẩu mở giúp lá hấp thu carbon dioxide cho quang hợp. Ngoài ra, chỉ số diệp lục tố trên cây xử lý NaCl kết hợp phun KNO 3 cũng được cải thiện hơn so với cây xử lý riêng lẻ NaCl 8 và 16 g/L ( Table 2 ), từ đó có thể làm chậm quá trình lão suy ở lá. Kết quả thí nghiệm này cũng đã cho thấy cường độ quang hợp được cải thiện, mặc dù vẫn còn thấp hơn so với cây đối chứng (NaCl 0 g/L, không phun KNO 3 ), nhưng quang hợp vẫn mạnh hơn so với cây chỉ được xử lý riêng lẻ NaCl ở cùng nồng độ. Bên cạnh đó, việc xử lý phun KNO 3 làm tăng hàm lượng proline tích lũy ở lá và rễ giúp cây chống chịu tốt hơn với điều kiện stress muối 18 . Hàm lượng proline tích lũy tăng cao ở lá và rễ trên cây xử lý kết hợp NaCl 8 hoặc 16 g/L và phun KNO 3 10 g/ L ( Table 2 ), proline tích lũy nhiều hơn các acid amin khác và điều này được sử dụng như một chỉ thị sinh hóa để đánh giá khả năng chống lại với điều kiện stress nước (do stress muối gây ra) 13 . Kết quả nghiên cứu trên cây lúa cũng tương tự, khi phun KNO 3 đã làm tăng tích lũy proline, cải thiện các chỉ tiêu sinh lý và từ đó giúp cây chống chịu với mặn 18 .
KẾT LUẬN
NaCl nồng độ 4‒16 g/L gây ra stress muối và stress thẩm thấu làm giảm sự tăng trưởng của cây chanh không hạt chiết nhánh sau 2 tuần xử lý. Việc tăng nồng độ muối ở trong đất đã làm tăng sự rụng lá, làm giảm sự tạo mới của lá, giảm trọng lượng tươi, giảm chỉ số diệp lục tố, giảm độ mở khẩu, giảm cường độ quang hợp theo thời gian. Stress muối đã thúc đẩy sự hóa lignin trong các tế bào nhu mô vỏ rễ ở cây. Hoạt tính ABA tăng ở lá tương ứng với sự đóng khí khẩu. Đồng thời, hoạt tính của IAA tăng cao, hoạt tính GA giảm dẫn đến đến sự tăng lão suy ở lá, đặc biệt là giảm sự tạo mới và kéo dài rễ.
Xử lý tưới nước rửa mặn giúp cây chanh ở nồng độ NaCl 4 và 8 g/L nồng độ NaCl 12 và 16 g/L chết hoàn toàn sau 6 tuần tưới nước phục hồi. Xử lý kết hợp phun KNO 3 10 g/L giúp tăng hấp thu K + ở lá, làm mở khí khẩu cùng với sự tăng tích lũy proline ở lá và rễ giúp cân bằng áp suất thẩm thấu từ đó làm tăng khả năng hấp thu nước theo dòng mạch lên lá.
DANH MỤC CÁC CH Ữ VIẾT TẮT
ABA: Abscisic acid
CCI: Chlorophyll Content Index (Chỉ số diệp lục tố)
IAA: Indoleacetic acid (auxin)
ĐHTTTV: Điều hòa tăng trưởng thực vật
GA: Gibberellic acid (gibberellin)
KNO 3 : Potassium nitrate
NaCl: Sodium chloride
XUNG ĐỘT LỢI ÍCH
Các tác giả đồng ý không có xung đột nào liên quan đến kết quả đã công bố
ĐÓNG GÓP CỦA CÁC TÁC GIẢ
Huỳnh Chí Hiếu thực hiện các thí nghiệm, thu thập, xử lý và viết bản thảo
Trần Thị Thanh Hiền định hướng, lên kế hoạch nghiên cứu, hoàn chỉnh bản thảo
Đỗ Thường Kiệt thảo luận các kết quả
References
- Carolina NR, Lucia MJ, Renata BF, Antonio GS, Edgar O. Valor nutricional de Lima ácida (Citrus latifolia T.), cv. Thaiti. Food Sci Technol. 2011;31. . ;:. Google Scholar
- Viện Khoa học thủy lợi miền Nam 2017. Dự báo xâm nhập mặn tại các cửa sông vùng ven biển đồng bằng Sông Cửu Long và đề xuất giải pháp chống hạn, Thành phố Hồ Chí Minh. . ;:. Google Scholar
- Barrosy J, Serk H, Granlundz H, Pesquet E. The cell biology of lignification in higher plants. Ann Bot. 2005;115:1053-74. . ;:. PubMed Google Scholar
- Deinlein U, Stephan AB, Horie T, Luo W, Xu G, Schroeder JI. Plant salt-tolerance mechanisms. Trends Plant Sci. 2014;19(6):371-9. . ;:. PubMed Google Scholar
- Hose E, Clarkson DT, Steudle E, Schreiber L, Hartung W. The exodermis: A variable apoplastic barrier. J Exp Bot. 2001;52(365):2245-64. . ;:. PubMed Google Scholar
- Hussain S, Luro F, Costantino G, Ollitrault P, Morillon R. Physiological analysis of salt stress behavior of citrus species and genera: low chloride accumulation as an indicator of salt tolerance. S Afr J Bot. 2012;81:103-12. . ;:. Google Scholar
- Mostajeran A, Rahimi-Eichi V. Drought stress effects on root anatomical characteristics of rice cultivars (Oryza sativa L.). Pak J Biol Sci. 2008;11(18):2173-83. . ;:. PubMed Google Scholar
- Việt BT. Sinh lý thực vật đại cương, Nxb Đại học quốc gia Thành phố Hồ Chí Minh, Thành phố Hồ Chí Minh. 2016;2016. . ;:. Google Scholar
- Ayala-Astorga GI, Alcaraz-Meléndez L. Salinity effects on protein content, lipid peroxidation, pigments, and proline in Paulownia imperialis and Paulownia foetune grown in vitro. Electron J Biotechnol. 2010;13(5):0. . ;:. Google Scholar
- Mohammad RA. Effect of salinity stress on growth, sugar content, pigments and enzyme activity of rice. Int J Bot. 2011;7(1):73-81. . ;:. Google Scholar
- Stoeva N, Kaymakanova M. Effect of salt stress on the growth and photosynthesis rate on bean plant (Phaseolus vulgaris L.). J Cent Eur Agric. 2008;9(3):385-92. . ;:. Google Scholar
- Negrao S, Courtois B, Ahmadi N, Abreu I, Saibo N, Oliveira MM. Recent updates on salinity stress in rice: from physiological to moleculer responses. Plant Sci. 2013;30(4):329-77. . ;:. Google Scholar
- Fahad S, Hussain S, Matloob A, Khan FA, Khaliq A, Saud S et al. Phytohormones and plant responses to salinity stress: a review. Plant Growth Regul. 2015;75(2):391-404. . ;:. Google Scholar
- Saeedipour S. Salinity tolerance of rice lines related to endogenous abscisic acid (ABA) level synthesis under stress. Afr J Plant Sci. 2011;5(11):628-33. . ;:. Google Scholar
- Hamayun M, Khan SA, Khan AL, Shinwari ZK, Hussain J. Effect of salt stress on growth attributes and endogenous growth hormones of soybean cultivar Hwangkeumkong. Pak J Bot. 2010;45(5):3103-12. . ;:. Google Scholar
- Ryu H, Cho Y. Plant hormones in salt stress tolerance. J Plant Biol. 2015;58(3):147-55. . ;:. Google Scholar
- Moore CA, Bowen HC, Scrase-Field S, Knight MR, White PJ. The deposition of suberin lamellae determines the magnitude of cytosolic Ca2+ elevations in the root endodermal cells subjected to cooling. Plant J. 2002;30(4):457-65. . ;:. PubMed Google Scholar
- Bo NV. Ảnh hưởng của KNO3, brassinosteroid và CaO lên sinh trưởng của cây lúa dưới điều kiện tưới mặn, Tạp chí Khoa học trường Đại học Cần Thơ, Số chuyên đề: Nông nghiệp. TR. 2014;2014(3):15-22. . ;:. Google Scholar





